PLANT HORMONES:
Introduction
By convention hormone are said to be a
substances whose site of synthesis and site of action are different; the two
events are separated by space and time. Hormones are known to elicit specific
responses. Charles Darwin first demonstrated the existence of such substances
in plants. Who in his own inimitable way explained growth of plant tips,
respond to light and exhibits photo induced curvature movements. Since then
botanists all over the world made studies in unraveling the mysteries of
diffusible substances called hormones which control growth and development of
the plant body.
Discovery of Plant Hormones:
Darwin used canary grass coleoptile tips to demonstrate the sensitiveness of the stem apex to light mediated curvature movements. Later Boysen Jensen used Avena coleoptile tips to demonstrate the presence of plant hormones. A gelatin block was placed on the decapitated coleoptile tip, then the tip was replaced over the gelatin block and the tip was illuminated from one direction. In response to light, the stem tip bent towards the light source. This was explained as due to the downward movement of some substance from the tip through gelatin block down wards. And the substance was considered as the cause for growth curvature.
Paal. A, on the other hand, placed the cut coleoptile tip asymmetrically over the decapitated coleoptile segments and placed the seedlings in dark. After few hours, he observed the curvature of the stem tip away from the side at which apical tip was placed. Pal’s experiment further demonstrated the presence of some kind of growth promoting substances in coleoptile tip, from which the substance was able to diffuse and bring about growth on one side hence the curvature. What ones people called diffusible substances have been restated as transportable substances for there are carriers in the cell membranes which do the functions? These are carriers specific.
F.W. Went collected growth promoting substances by placing the coleoptile tips on the square agar blocks. By placing such loaded agar blocks asymmetrically on the decapitated coleoptile tips in dark, showed the growth curvature movements. Perusing the above methods he established quantitative bioassays. The bioassays explain and correlate quantitative relationship between the amount of hormone applied and the magnitude curvature as a response.


Both meristems are composed of undifferentiated and dividing cells that deliver new cells to the growing structures. Small populations of stem cells (SC) within the meristems are essential for the self-perpetuating capacity of meristems. Following asymmetric divisions, the stem cells form daughter cells with distinct cell fates. One daughter cell will maintain stem-cell fates, whereas the other will receive instructions to differentiate into a specific cell type. However, the final differentiation is postponed and will only occur after several rounds of symmetric divisions that give rise to growing cell files (arrows). In the shoot apex, leaf primordia (LP), also composed of dividing cells, are formed in close proximity to the SAM. In the root, no new organs are formed near the RAM and the entire structure is covered by a protective root cap (RC). At the apex–cap boundary of the root, a small group of non-dividing cells collectively known as the quiescent centre (QC) is present. Signals emitted by these cells maintain the immediate surrounding cells in a stem-cell state (blocking lines) and prevent them from differentiation. http://www.nature.com/

www.lhr.ueb.cas.czwww.cell.com
It is at this juncture of time biologists and chemists started identifying the chemical component that is responsible for growth promoting activity. To their surprise, they found the human urine as a rich source for the said hormone. Kogl and Haagen Smith starting with 33 gallons of urine, extracted 40 mg of the active principle in the form of crystalline powder which showed 50,000 fold grater activity. First they called this substance as Auxin-A. Using the same extraction procedures they isolated another active substance from corn germ oil and called it as Auxin-B. Not satisfied with their purification methods, they used charcoal adsorption column chromatographic procedures for isolating a pure form of growth substance. The substances obtained from this method were called Heteroauxins. Later heteroauxins were identified as Indole Acetic Acid. But this substance was known as a chemical to them for it was already identified by E & H Salkowaski. However, Salkowski’s did not know about the properties of IAA as growth hormone.
The discovery of Indole acetic acid as the plant growth hormone gave impetus to plant physiologists. As a result, new hormones were discovered from different plant sources. Their site of synthesis, chemical structure, site of action and their physiological and morphological effects have been studied in detail. So far, five phytohormones have been identified from different parts of the plant body; most importantly all the hormones can be detected in the same plants. They are Indole Acetic Acid (IAA), Gibberellic acid or Gibberellins (GA), Cytokinin, Abscisic acid (Abscisin or ABA) and Ethylene. Simultaneously a host of synthetic compounds have been developed which stimulate plant hormones in many respects. The figure below shows hormone synthesis pathway.

Pathways of few hormone’ synthesis; www.devids.net
Phytohormones and Plants’ responses:
Investigation on the effects of hormones on plants has revealed that the hormones elicit a wide array of responses in different types of tissues in the same plant body. Physiologists have realized that the responses to the hormonal treatments depend upon the kind of tissue and the physiological state of the tissue. For example, in a developing stem segment, in response to GA3, the internodes elongate considerably but the same hormone in maize grains elicit the synthesis of Alfa amylase enzymes is aleurone cells. Thus the specific response to a particular hormone depends upon the inbuilt potentiality of the said tissues; this behavior is because of its previous developmental programs. It should also be remembered that the specialized hormones found in animal systems have no counterparts in plants with respect to the target cells and specific functions. The Auxin which induces the growth in one part of the plant body, fails to bring about the same effect in the other part, but it may have different effects like apical dominance, new root formation or parthenocarpy, etc., at different site. While GA is known to bring about the gene activation in aleurone cells leading to the synthesis of alfa amylase, the same hormone acts on rosette shaped Hyoscyamus plant and induces bolting and flowering. But in pea plants, it overcomes genetic dwarfism. The above observations suggest that each and every phytohormone elicit more than one response in the same plant body but at different sites. Furthermore as all hormones are synthesized at different sites within the same plant body the said hormones interact with each other and control the growth and development.
Hormonal Interplay:
Although Haberlandt considered the tissue and organ culture under sterile conditions as a theoretical possibility, Kattle in Germany and Robbin in USA have succeeded in developing methods to culture tissues in a defined media. With the advent of tissue culture techniques, studies on experimental morphogenesis progressed in leaps and bounds in the past 45 years or so.
Using tissue culture methods, it is possible to treat the tissue with one or the other hormone at will. Use of phytohormones in tissue cultures, have revealed that though a single hormones has a specific effect, two or more hormones together at different concentrations, elicit different responses in tissue from. In the presence of two different hormones, the effects may be promotive, synergistic or antagonistic where one may modify the activity of the other. Such reactions are referred to as Hormonal interplay.
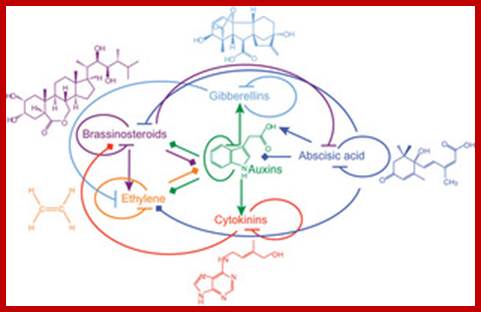
Plant hormones play a major role in plant growth and development. They affect similar processes but, paradoxically, their signaling pathways act nonredundantly. Hormone signals are integrated at the gene-network level rather than by cross-talk during signal transduction. In contrast to hormone-hormone integration, recent data suggest that light and plant hormone pathways share common signaling components, which allows photoreceptors to influence the growth program. We propose a role for the plant hormone auxin as an integrator of the activities of multiple plant hormones to control plant growth in response to the environment.; http://www.nature.com/structuraland Molecular Biology;


Network of Hormone Effects on Hormone Metabolism: Genes assigned to hormone biosynthetic pathways by GO annotation were identified within lists of hormone-responsive genes. Lines with arrowheads represent up regulation of hormone biosynthetic genes or down regulation of genes involved in hormone inactivation. Blocked arrows represent down regulation of genes involved in hormone biosynthesis or up regulation of genes involved in inactivation of a hormone. Diamond arrowheads indicate changes in gene expression with ambiguous outcomes (i.e., genes affected include those linked to both increased and decreased hormone levels). http://www.sciencedirect.com/

Schematic interaction between GA, ABA and Ethylene signaling pathways in regulation of seed dormancy and germination; http://www.ssedbiology.de
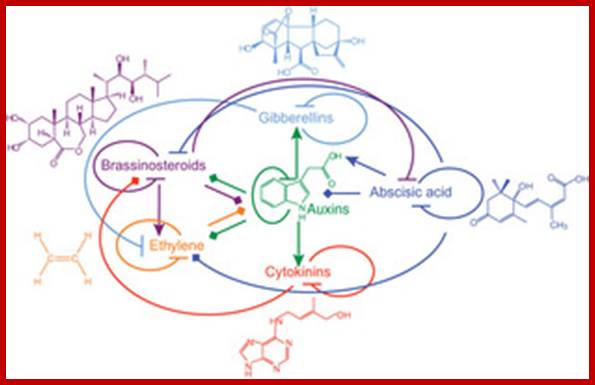
Phytohormone structure and functional interactions: Yvon Jaillais
& Joanne Chory; http://www.nature.com/
‘Lines with arrowheads, up regulation of hormone biosynthetic genes or down regulation of genes involved in hormone inactivation; blocked arrows, down regulation of genes involved in hormone biosynthesis or upregulation of genes involved in their function. Plant hormones play a major role in plant growth and development. They affect similar processes but, paradoxically, their signaling pathways act nonredundantly. Hormone signals are integrated at the gene-network level rather than by cross-talk during signal transduction. In contrast to hormone-hormone integration, recent data suggest that light and plant hormone pathways share common signaling components, which allows photoreceptors to influence the growth program. We propose a role for the plant hormone auxin as an integrator of the activities of multiple plant hormones to control plant growth in response to the environment’ Yvon Jaolais and Joan Chory.

Plants using CO2 and sunlight and water produce a variety of metabolites required for the plant; www. nptel.ac.in

http://biology.tutorvista.com/

Callus tissue can be grown from tobacco pith cells or from any other plants and the same can be maintained in a defined nutrient medium in the presence of both IAA and cytokinins at particular concentrations. The same callus can be induced for organogenesis by changing the relative concentrations of IAA to that of cytokinins. In the presence of higher amounts of auxin to cytokinins, callus generates roots. On the contrary if the concentration of cytokinin is higher in relation to auxins, the callus cells produce shoots only. If the concentration of the two said hormones is balanced, the callus induces the formation of both roots and shoots.
The above mentioned observations suggest that the hormonal interplay has a significant role in organogenesis. Similarly, GA3 promotes callus growth but the same hormone inhibits auxin-cytokinin induced shoot formation. The above said hormonal effects are not just direct effects, but they also influence the levels of other hormones either by activating the synthesis of a particular hormone or by inhibiting the synthesis of it.
Various studies on hormonal effects show that the endogenous levels of auxin in plant tissues is elevated by the applications of GA, Cytokinins or both. While auxin induces the synthesis of ethylene which in turn induces the formation of ABA, which on the contrary enhances the levels of ethylene? Thus, they show both cooperative and promotive effects on each other. It is also known that ethylene and ABA together bring down the levels of the auxin. This effect can be overcome by the addition of cytokinins, for cytokinins are capable of bringing down the levels of ABA through the increased levels of GA biosynthesis. The close relationship and interplay between GA, auxins, cytokinins, ABA and ethylene, exhibits an excellent feedback control mechanism. However, understanding of hormonal interaction at the level of gene expression and their product is very important in interpreting the hormonal interplay and effects.