PHYSIOLOGY OF PLANT MOVEMENTS
Higher plants, being fixed to soil cannot move from place to place. But within the plant body various protoplasmic components are in constant motion, ex., movement of water, minerals, food etc. But certain parts of the plant body in response to external stimuli, exhibit physical displacement called movement. Lower unicellular plants also show movement from place to place. Such movements may be autonomic or induced.
The movement of plant structures in response to stimuli is very interesting. The stimuli might be in the form of light, touch, chemicals, temperature, gravity or water. Thus the agency or factor that causes movement is called stimulus. Not all parts of the plant act as sites of perception to stimuli. Only certain regions or structures are capable of receiving the stimuli and such structures or organs are called perception site. The site of perception need not be the structure that responds to movement. In fact, in many cases the site of response and the site of perception are different. Nonetheless, the stimulus has to cross through the plasma membrane of the cell or cells found in such structures that receives the stimulus. Plasma membrane has all the inbuilt components and the potentiality to receive stimulus and transmit it into intracellular milieu for proper response. Examples; root movement towards water and minerals; carnivorous plants organs show movements in response to Insects, pollen tubes move (growth) towards egg in the ovary along the style and stomata open and close in response to light and moisture.
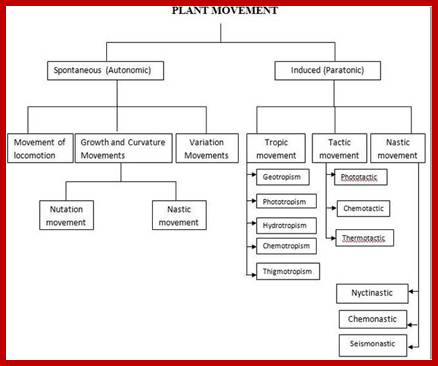
Types of movements (some): Circumnutation- rotary or helical ex. Tendrils. Tropic movements; curvature movements; Photoptropism; stem terminals move toward light – can be positive or negative; Geotropism or Gravitropism-response to gravity; Chemotropism: response to chemical stimulus; Heliotropism: towards light; Rheotropism; movement of curvature-plant organs; Thermotropism movement in response to touch and Traumatropism: in response to injury.
Nastic movements- independent direction of stimulus, where the direction is predetermined: Epinasty, Hyponasty, Hydronasty, Nyctinasty, Seismonasty, Thigmonasty or hyponasty.
There is a time lag between the time at which the stimulus is applied and the time at which the response begins. This time is called reaction time. It may vary and depends upon the intensity of stimulus and the kind of response. If the stimulus is weak, there may not be any response at all, but if the stimulus is adequate or in right quantity the response is positive. The time required to cause the proper stimulus is called presentation time.
Once the plant body responds to the stimulus say sleeping movement (change in the position direction), structures involved always come back to their original position. This process is called recovery and the time required is referred to as relaxation time, which again varies from species to species. If the stimulus is provided repeatedly the receiver structures do not respond with the same intensity as the first instance, but it slows down. This effect is due to fatigue. Such behavior is probably due to the loss of same material components required for the response. If the stimulation is continued at increased frequency, the plant organs do not respond and behave as if they are dead structures. Such a state is called Tetanus or extreme fatigue. Now it is believed that stimulation (quantity) causes irritation at the site of perception and the products of irritation are then transported to the site of response, where the structures respond and perform movement. All these events initiate with signal, and the receptor that receives the signal becomes active and induces signal transduction pathway in the cell cells. There will be a cascade of events that finally leads to the response.
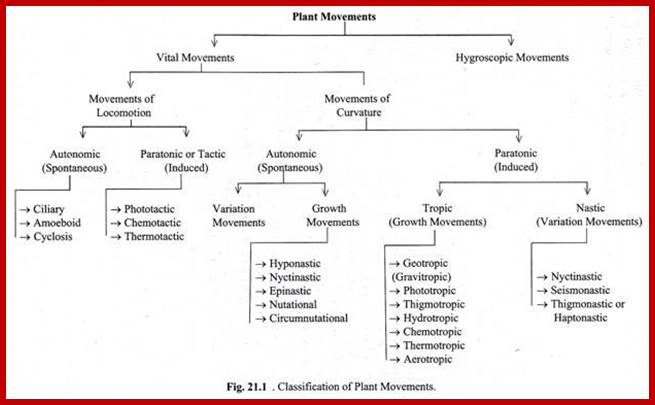
Plants endowed with different structural adaptation and different potentialities exhibit different types of movement either voluntarily or involuntarily. Based on the behavior as pattern, movement’s stimulus, the plant movements have been grouped into physical movements and vital movements.
Physical Movements:
The structures involved in this type of movement are mostly dead. Xerochasy: Structures like the wall of fruits, sporangia, capsules, have differential wall thickenings. During dry weather conditions, they loose water to atmosphere. Because of this, the thick walls contract as a result they break open along with the line of dehiscence, where the cell walls are thin and susceptible. Hydrochasy: Certain structures, made up of hydrophilic substances, are capable of imbibing water as well. Due to imbibition of water they swell. Due to imbibition of water peristomial teeth in Moss capsules, elators in equisetum, etc. show movements. In fact such movements help in the dispersal of spores.
Vital Movements:
Plant movements due to the activity of living structures are called vital movements. They are further classified into different kinds. In some plants, particularly unicellular algae, the entire cell moves from place to place or from one position to the other or the protoplasm by itself shows continuous flux by physical displacement. In others, where the plant body is fixed in the soil, certain structures show bending or curvature movements. Furthermore, some of the movements are auto regulated and propelled by innate mechanisms but other movements are induced by stimuli.
In this case the entire living cell is involved either in the movement of protoplasm or the entire body of the plant cell from one place to another. These movements may be autonomous and induced.
Autonomous: Protoplasmic Streaming:
Protoplasm is not a static fluid. With all its complicated structures the entire protoplasmic fluid is in constant sweeping motions. These streaming movements can be observed under high resolution light microscopes. For example, in the cells of staminal hairs of tradescantia, the streaming is localized and in each of these areas the direction is clockwise or anticlockwise. Tiny particulates are seen swept along with the stream in a particular direction and particular directed path. Such compartmentalized movements are known as Cyclosis. But in Elodea and other plants, the protoplasm shows uniform movement, but in one direction. One can observe chloroplasts movement along the cytoplasmic streaming. Such movements are called Rotational Movements.
Protoplasmic streaming is due to the activity of contractile proteins (Actin-Myosin) found associated with other microtubule but microtubules provide direction and actin-myosin network within the cytoplasm is responsible for movement. Charal cells are useful for they show unidirectional flow; at periphery the movement is fast than towards the vacuole. It is an active process microtubules play an important role in such intra cellular movements. These structural elements contain motor proteins and they perform the movements. The bulk of endoplasm and cell sap move passively. The rate of streaming was found to be 100um/sec. Active protoplasmic streaming has been observed in developing pollen tubes. Cytoplasmic organelles enmeshed in ER display Brownian movement while streaming. The ER membranes slide along Actin fibers. The energy required for this process is derived from ATP molecules. These movements help in the even distribution of chemical components. And the protoplasmic movement across the plasmodesmata brings about the transportation of materials from one cell to another. Auxin has been found to accelerate the rate of protoplasmic streaming. Addition of colchicine and cytochalasin B totally inhibits the movement, thereby indicating the involvement of microtubules and microfilaments. Respiratory poisons like DPN, KCN, also inhibit the movement, thus suggesting that these are active movements. Even amoebae show such movements. For that matter all living cells exhibit autonomous movements.
Paratonic movements:
These movements are stimulated by external agents like light, chemical heat, etc.
Hence, they are called taxis or tactic movements.

www.tutorvista.com
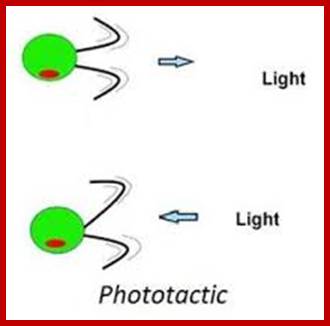
Phototactic Movement: Unicellular algae suspended in a test tube moves towards the light source to obtain solar energy for photosynthesis. If the light is very intense, they move away from light; this may be due to the raise in temperature. The movement in these cases is due to loco motor structures like flagella or cilia. The beating of these structures propels the cells towards light. The flagellar activity utilizes ATP. Hence these movements are active.
Biology4disc.weebly.com
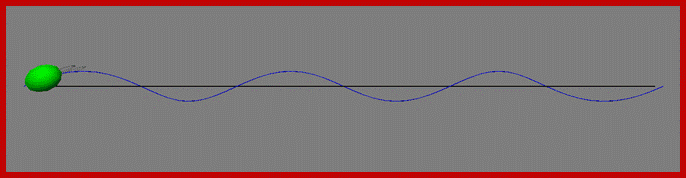
Phototaxis system of chlamydomonas , the cell swims , rotates along a helical path; and stirs with the beating of a pair of Celia: Light signals alters transmembrane electric current that results in transmembrane electric field changes in Celia that in turn differentially control the response of two Celia steering the cell; http //fosterlab.syr.edu/
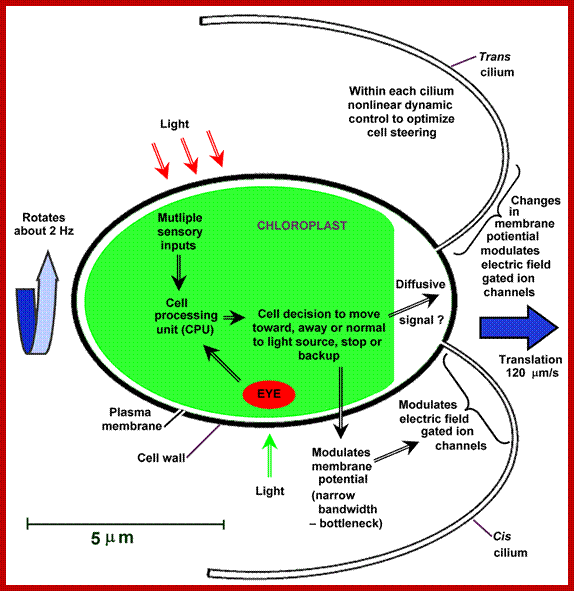
Intracellular processing determines the celiary behaviour such as the direction of phototaxis and how the tracking of light source is optimized ; celiary beating required ATP energy in the cell; pigment rhodopsin activation and photo-movement is involved. http //fosterlab.syr.edu/
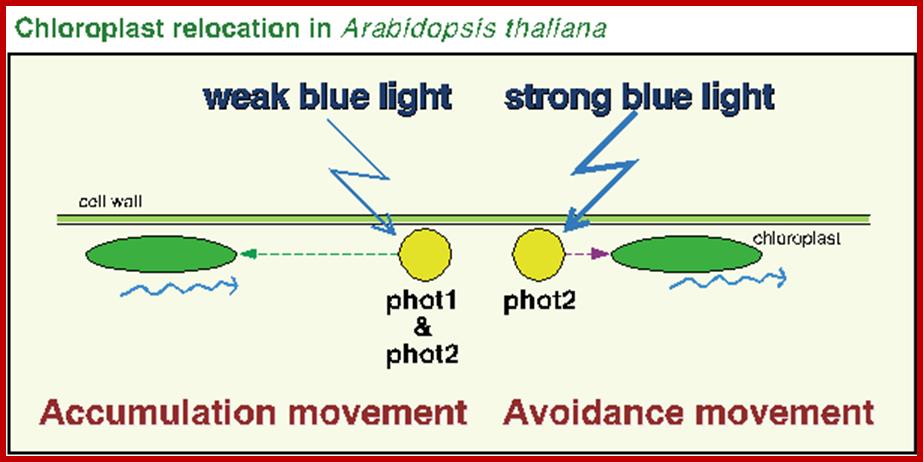
Chemotactic Movement; Certain flagellated or non-flagellated bacteria move towards the source of food by lashing their flagella or by tumbling. In these cases, the stimulus is chemicals. Similarly, the movement of spermatozoids in lower organisms like Bryophytes is directional, because the chemicals released by mature archegonia provide the chemo stimulus. Sensing the chemicals, the flagellated spermatozoids swim towards archegonia and enter the neck canal and bring about fertilization. Pollen tube grows toward ovules along the style.
Thermotactic: Again lower organisms sense the temperature and move towards the compatible temperature or move away if the temperature is incompatible. Sensing the change in the temperature by the plants is autonomic but the cells always show directional movements.
All the above mentioned movements involve signal transduction pathway. Elucidation of these pathways at molecular level is necessary and exciting.
Movements of Curvature:
Plants with their fixed plant body cannot move from place to place, but certain structures show bending movements which may be directional or non-directional or they may be autonomous or induced. However, some of the plant movements are due to the growth of the cells or due to change in the turgidity of the cells. Based on the above features movement of curvature has been further classified such as:
Autonomic Growth Movements:
Nutatory: Plant structures like tendrils and stem tips exhibit differential growth alternately. This results either in sideward or circular movements. Certain sub cylindrical stems (flat or angular) due to differential growth at the sides show zig zag movement. On the other hand, cylindrical tendrils in search of getting a hold on to a substratum; waves of growth activity takes place around the tendril. That is why it appears as if it is moving in circles. Such type of movements is called circumnutatory movement. The growth activity of the above said structures is in-built and they show a rhythmic pattern. Whether the hormonal fluxes are responsible for this type of growth movement or any other innate mechanism involved is not known. But the treatment of these stems with ABA inhibits the movements, indicating that these are phytohormone mediated.
Ephemeral Movements:
During the development of leaves and floral organs, the growth pattern determines the growth direction of the structures. For example, leaves expand laterally by continuous radial divisions and expansions. Similarly sepals and petals because of continuous growth of cells at the base on the inner surface make the flower open. The hypocotyl hook of the bean seed straightens up because of one sided growth due to expansion cells on that side. Such growth movements are called Ephemeral. Once the movement reaches, a particular stage the bending movement stops.
Autonomic Turgour Movement:
Desmodium gyrans (Indian telegraphic plant) and Eleiotis sorria have trifoliate compound leaves. In the former case, central leaflet is larger and straight. But the lateral two leaflets are smaller which show regular upward and downward movements. Such movements are observed only during daytimes but not at night. Such rhythmic gyratory movements require about 5-8 minutes for the completion of one cycle. This is due to autonomic turgour changes in the cells found in the swollen pulvinus of the leaflets. How light brings about turgour changes or is it due to phytochrome mediated responses or hormonal responses is not clear.
Paratonic Nastic Movements:
Paratonic movements are induced by external stimuli. Whatever may be the point or direction of stimulus applied the movement of the plant structures is already predetermined and they exhibit movement only in one direction. Most of the movements are turgour movements, but growth movements are not uncommon. For example, the opening of hypocotyl hook of germinating bean seedling, opening of circinately coiled leaves of ferns and cycas are the examples of growth mediated nastic movements. Differential distribution of growth hormone in the adaxial (dorsal) surfaces of the leaves; it is this that is mainly responsible for such growth movements. These movements are permanent and they do not show temporary day and night fluctuations.
It is due to difference in the rate of growth on two opposite surfaces of the plant organ due to faster growth on the upper surface of the organ; Epinasty- growth at the lower surface and hyponasty due to growth below.

Nastic movement-hyponasty-http://biology4isc.weebly.com/
Photonasty: Many leguminous plants, with their pulvinous bases, show characteristic sleeping movements and they show a rhythmic pattern of opening of leaf-lets in the day and closing in the evening with a precision of a clock. In fact, such movements are attributed to circadian rhythm operated by an inbuilt biological clock.
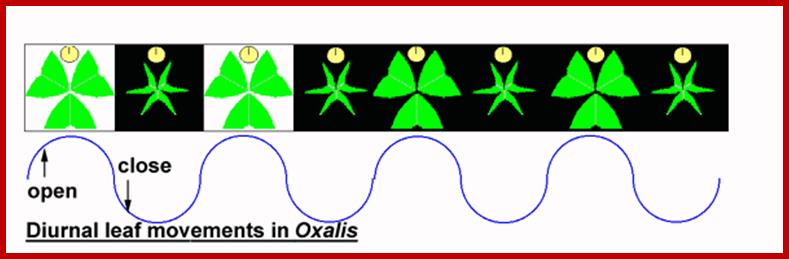
www.s10.lite.msu.edu
The photonastic mechanism has been explained on the basis if hormonal distribution in the cells of the pulvinus. During daytimes, auxin is found in greater amount at the upper region of the pulvinus, because of this the cells become more turgid and leaf lets open. The same process is reversed during night because of the redistribution of auxin to lower side which causes folding by the way of turgour changes.
Investigations into such movements have shown that these are phytochrome mediated, because red and far red lights are very effective in opening and folding of leaflets. Phytochrome being an excitable molecule in response to light it is known to bring about changes in the permeability of membranes. Thus the turgour movements bring about so called sleeping movements. Added to this the change in the permeability also involves the efflux and influx of K+ ions. The loading or unloading of potassium ions causes increased or decreased DPD which acts as the driving force for the entry or the exit of water which results is turgour movements.
Chemonasty: Drosera, Venus fly trap and such insectivorous plants have devised mechanisms to trap insects to obtain nitrogenous compounds. These plants are found growing in boggy areas and they are incapable of utilizing soil nitrogen.

www.mentalfloss.com
The plant Drosera consists of leaves arranged in a rosette pattern. The leaf blades are flat and racket shaped. The upper surface is made up of colorful glands which secrete sticky juice, where you very high activity of golgi complex. There are a large number of sensitive tentacles spread out at the margins. The tentacles have a sensitive broad base and a terminal glandular globosely head which also secretes juice. In sunlight, these structures glisten like dew drops; hence the name sundew plant.
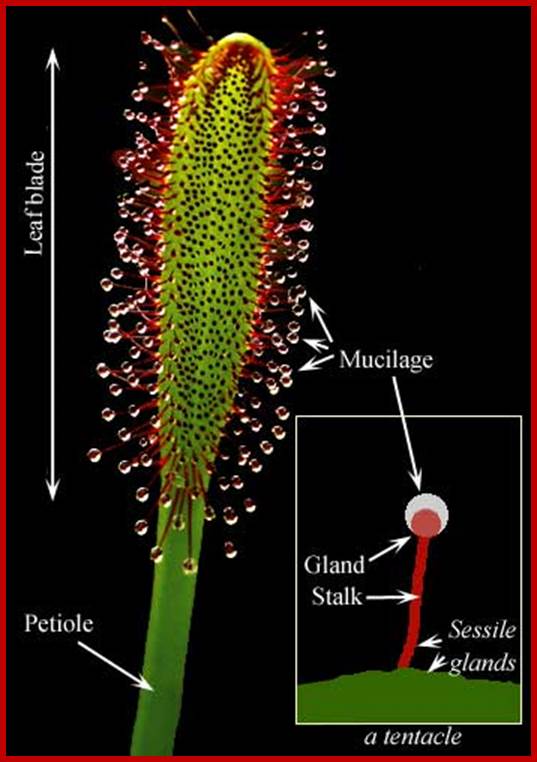
http://www.puchen.org/
Insects, mistaking the glistening juice for honey, settle down on these surfaces where their legs get stuck because of the sticky juice and they cannot escape. As insects have nitrogenous compounds in their body, they diffuse and sensitize the tentacles, because nitrogen compounds act as signals, in response, they immediately fold upon the insect and the same gets trapped. In this case nitrogenous compounds found in insects provide the stimulus. On the other hand, if a metal piece is placed, tentacles do not fold, instead if a piece of meat is dropped, the tentacles close immediately. However, after digestions, tentacles open slowly and this process takes few hours. Such chemonastic movements are found in utricularia, Venus fly trap, etc.
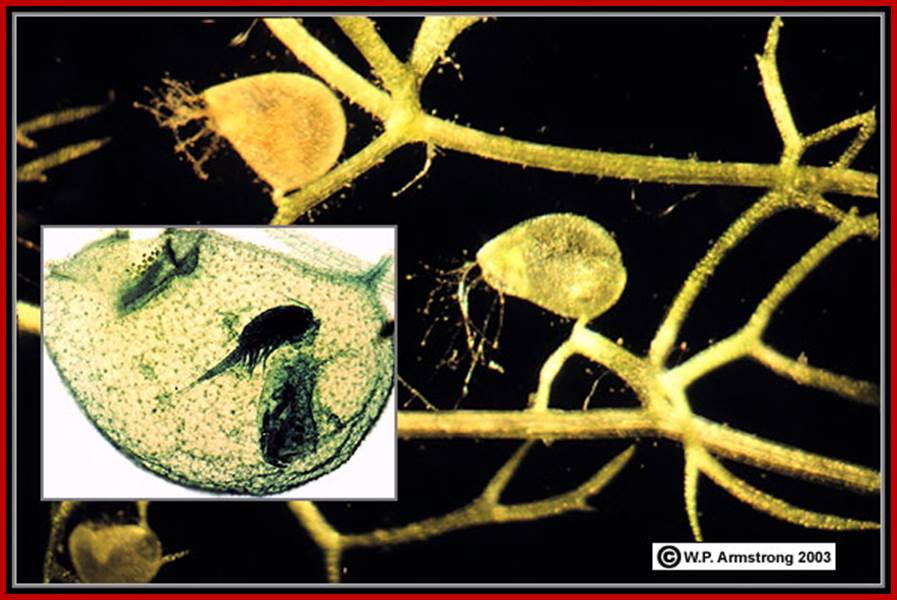
www.top-10-list.org
Thermonasty: Flowers in Tulips and Crocus are very sensitive to temperature variations. Accordingly the accessory whorls may be closed or open. These movements are known to be due to turgour changes rather than growth movements.
Siesmonasty: Mimosa pudica (Touch me not plant), Biophytum, Neptunia oleracea, Desmanthus are very sensitive to touch, rather shock generated by the touch. Mimosa pudica has pinnately compound leafless with a swollen pulvinus at the base every leaflet. Touching the leaves is believed to cause a seismic shock to the leaves and this stimulus is transmitted all along the rachis downwards and reaches the basal pulvinus of the leaf. The irritability caused, due to touch is actually transmitted through sieve tubes, because these are the only structures which are capable of transmitting the stimulus as fast as 1.5-20 cm/sec. The material basis for the transpiration stimulus has been found to be ABA and ABA mediated ions. As the ABA ions diffuse along the rachis, it also trans-diffuses into pulvinus of the leaflets.

www.prezi.com



Basal turgor movements; http://fabacademy.org/http://en.wikipedia.org/
The mechanism of upward movement of pinnules and downward bending of the entire leaf at the base is now considered as due to the activity of the motor cells found in the pulvinus. Anatomical studies indicate that at the lower region the pulvinus consist of thin walled parenchymatous cells, which are loosely arranged with lot of intercellular spaces. Cells also contain many contractile vacuoles. The cell membranes and vacuolar structures have contractile proteins. These vacuoles are loaded with K+ ions when the leaflets are open. These motor cells are highly sensitive and active. Such cellular organization is also found at the upper region of the pulvinus found at the base of the leaflets.
When the stimulatory hormone ABA, which is also called as stress hormone (released due to irritability) reaches these specialized cells, they are stimulated to contract, collapse and extrude water and K+ ions into intercellular spaces. This action brings about the collapsing of cells on this side and the leaf downwards. Similarly mechanism is involved in the folding of leaflets upwards. The recovery takes place within 5-10 minutes. The opening process takes place by active pumping in of K+ ions back into cells. The K/ATPase pumps are supposed to be found in the cell membranes. This active process requires energy. If the production of ATP is inhibited by respiratory inhibitors like DNP or KCN recovery does not take place. If plants are kept is continuous dark, they fail to produce any siesmonastic responses, which suggest the siesmonastic movements are ATP dependent active processes.
Paratonic Tropic Movements:
These movements are due to growth activity. The curvature movement is always directional, either towards the stimulus or away from the stimulus. Most of these are phytohormone mediated movements.
Phototropism: Phototropic movement is mostly exhibited by stem tips; for they always bend towards the light source. The action spectrum of the light has been found to be at blue light. The blue light is now known to bring about unequal distribution of active auxins than the total auxin content. The pigment for absorption of blue light has been found to be Cryptochrome. The older view of destruction of auxin and the lateral movements of auxins in response to light has been more or less ruled out. The difference in the quantity of active auxin brings about differential growth and also curvature. Details of this process have been explained elsewhere.
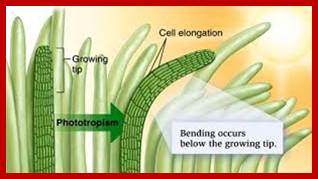
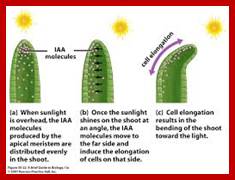
www.cannabisgrowguide.wordpress.com; www.alandalus.blogspot.com

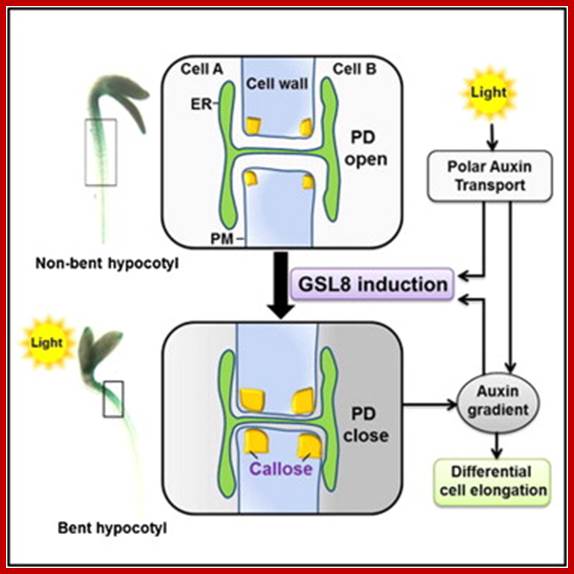
In plants, auxin functions as a master controller of development, pattern formation, morphogenesis, and tropic responses. A sophisticated transport system has evolved to allow the establishment of precise spatiotemporal auxin gradients that regulate specific developmental programs. A critical unresolved question relates to how these gradients can be maintained in the presence of open plasmodesmata that allow for symplasmic exchange of essential nutrients and signaling macromolecules. Here we addressed this conundrum using genetic, physiological, and cell biological approaches and identified the operation of an auxin-GSL8 feedback circuit that regulates the level of plasmodesmal-localized callose in order to locally downregulate symplasmic permeability during hypocotyl tropic response. This system likely involves a plasmodesmal switch that would prevent the dissipation of a forming gradient by auxin diffusion through the symplasm. This regulatory system may represent a mechanism by which auxin could also regulate symplasmic delivery of a wide range of signaling agents. http://www.cell.com/developmental

Phototropism of the sporangiophore (fruiting body) of the zygomycete, Phycomyces blakesleeanus (photograph by D.S. Dennison). www.uni-marburg.de
Phototropic movement in in one of the fungi called Phycomycs blacesleeanus
The apoprotein cry in association with flavins bring about photoptropism, it gets activated by absorbing blue light and autophosphorylation of ser/thr residues, that leads to transport of ions to one side and brings about bending movement.
Geotropism: Growth of roots towards soil and the stem away from the soil in response to gravitational force is an inbuilt mechanism endowed in stem tip and root tip of the plant. Curvature movement of roots towards gravitational is termed as positive geotropism and the movement of the stem away or against gravitational force is called negative geotropism. However in some plants, rhizomes and runners grow parallel to the surface of the soil. Such a response is called digeotropism. The obliquely growing stems show plagio geotropism (For details refer the Chapter Auxin).
The differential response of stem tip and root tip to the same gravitational force is due to their different innate structural and functional potentials. Though both structures are derived from the same embryonic cell and possess the same genetic potential they behave differently probably this is because of the programming of process driving the development of in such structures. The mechanism of geotropism has been explained on the assumption that differential responses are due to different concentrations of auxins. The concentration of auxin that promotes the growth of the stem apex inhibits the growth of the root tip. On the other hand, the concentration of auxin that is effective in the growth of the root tip is not adequate for the growth of the stem tip. This concept suggests that stem apex requires higher concentration of auxins for its growth and roots require low levels of auxins as optimal concentration for the normal growth.
If a seedling is placed on the soil horizontally, due to mass action of gravitational force, auxins move downwards in both stem apex and root apex. In the stem apex, as more and more of auxin accumulates at the lower surfaces the cells found in this region grow faster than the upper cells, thus the stem curls upwards. But in roots, as more of auxin accumulates at the lower region, the higher concentration inhibits the growth of cells found at the lower surface, but the cells at upper surface grow faster because of lower concentration of auxin. Furthermore, the root cap being the preceptor of gravitaropism produces ABA, which on translocation basipetally, reaches the lower cells by sheer gravity. With the accumulation of more and more of ABA at lower cells, the growth of these cells is further inhibited, but the cells at the upper region grow normally and thus bring about the downward growth curvature for roots.
Chemotropism: Pollen tubes and certain fungal hyphae exhibit chemotropic movements, because they grow towards organic nutrient rich media. In the case of pollen tubes, the direction of growth is dictated by the chemical gradient generated by the embryo sac. This chemical gradient greatly facilitates the growth movement of pollen tube towards the embryo sac, irrespective of the position of ovules in the ovary.
Hydrotropism: Growth of roots towards water is called hydrotropism. Whatever may be the positions of seedlings, as shown in the figure, the roots curl towards water. This positive growth curvature might be due to greater water potential, probably acts as the motive force for the growth of root tip. Stem and other structures are insensitive to water and they do not show any hydrotropic curvature movements.
Thigmotropism: Plants with weaker stem spread around and require support for their growth. Many of the climbers have developed hooks, pads and tendrils for getting a firm foot hold onto the substrate; which may be a rock or a wooden stick or any such hard structure. For example, in the case of tendrils of cucurbita and other species the terminal regions are soft, tender. The terminal region of the tendrils consists of a number of fine pits, which are highly sensitive to touch. When the tendril comes in contact with any supporting structures, the pits get stimulated and the same is transmitted a few millimeters basally. This results in the differential growth of the tendril on one side. This causes the curvature in the tendrils and finally they coil around the stem the supporting structure. The time required for such coiling after the stimulus is just 3 to 5 minutes or more. Once the initial reaction is sets in at the basal region, the tendrils coil round the support and thus draw the climber nearer to the support. It is suspected that auxins and ABA are involved in thigmotropic curvatures. ABA inhibits the growth of the cells in the region of the contact and auxins stimulate the growth of the cells on the opposite side and bring about eh growth curvature called ‘coiling’.

http://www.doeaccimphal.org.in/


Tendrils-Thigmotropism; https://www.quora.com

http://krivsky-thigmotropism.yolasite.com/