Physiology of Vernalization
Vernalization and Flowering
Light has a profound influence on plants and it performs many important biological processes like photosynthesis, phototropism, photorespiration, photoperiodism, etc. Influence of light on plants in flowering is very fascinating, but not all the plants respond and flower to photoperiodic treatments. Similar to light, the temperature also has significant effects on plant growth, dormancy and flowering. Most of the metabolic processes are regulated by the temperature that is prevailing in the environment. But the effect of temperature in inducing the development of reproductive organs is fascinating.
As mentioned earlier, different plants have different cycles of vegetative growth, flowering and fruiting. However, some biennials which produce vegetative structures in one season and induce flowering in the flowering season only after they are exposed to prolonged winter or cold treatment. Interestingly, such cold requiring plants also need proper photoperiodic treatment for flowerng. Such biennial plants can be made to flower in the same season by subjecting them to cold treatment. Hence the process of acquiring the ability or capacity to accelerate the process of flowering, in response to cold treatment is referred to as Vernalization. In Soviet Union, the same phenomenon is called Yerrovization or Lysenkoism. Ever since Gassner (USA) discovered this unique phenomenon, a large number of plants which required vernalization have been identified and detailed studies have been made about the behavior of such plants. After the Second World War, under the supervision of Lysenkov in USSR extensive work has been done on vernalization and the same was exploited in the field of agriculture.
The winter rye called Secale cereale or Petkus rye is a binnial plant which requires cold treatment for successful cultivation as one season crop. The grains of these plants are known for their hardiness and quality for the purpose of milling and baking.
In fact, farmers used to cultivate this variety after subjecting the water imbibed grains to cold treatment and growing them in the spring and harvesting in the same in summer. After Klippert, who reported this phenomenon, extensive research work has been done in this field. Among many plants, Petkus rye (short day plants) was the first to be used for experimentation. The other examples are Hyoscyamus niger (long day plant) Triticum aestivatum (CV winter wheat), Lunaria bienensis, Arabidopsis thalliana, Lolium perennial, Beta Vulgaris, Brassica oleracea, and others.
Age and Site of Vernalization:
Vernalization through cold treatment is very effective at the seed stage or seedling stage. In some cereals, even the embroyes can be successfully vernalized. However, in many cold recurring species, vernalization is not effective until and unless the plant possess at least few leaves. The requirement of few leaves for effective vernalization is called ‘Ripeness to Flowering’. This suggests that plants need certain degree of photosynthetic inputs to obtain and to respond to cold treatment. Additional support to this view comes from an experiment on the embroyes of petkus rye. The embryos are separated from the endosperm and then if the embryo alone is subjected to cold treatment, embryos fail to be vernalized. But if such complete embryos are subjected to cold treatment along with carbohydrates like sucrose solution or endospermous tissue, vernalization is very effective. Probably the role of carbohydrates in vernalization is to supply some energy. Nevertheless, the most sensitive site which acts as the perceptive organ is the meristematic region of the shoot apex. Even leaves which act as the sites should have certain amount of meristematic tissues in them. But how does the cold treatment brings about this effect in such active cells is not known.
Temperature Effect:
For the normal growth and development, every plant requires on optimum temperature. But for vernalization the optimum temperature required is 3 0C to 17 0C, which varies depending upon the species involved. Even the duration of treatment varies from species to species. Individual requirements have to be determined independently by experimentation. In petkus rye the most effective range of temperature is 3 0C to 7 0C. But for Hyoscyamus niger 3 0C -17 0C degree is optimal. However the efficiency of cold treatment in bringing about vernalization is determined by the number of days shortened between germination and flowering stage.

Thinking about the dependence of human nutrition on fruit, seeds and their products leads to a realization of the importance of the regulation of flowering. Rightly then, the transition from the vegetative to the flowering state continues to attract attention. However, the floral transition is not just about forming flowers, as is illustrated by the work of Cookson et al. (Montpellier, France, pp. 703�711). Their subject of study was the favorite �model� long-day plant, Arabidopsis thaliana. Plants were grown under five different day-length regimes; in all other respects treatments were identical. We concentrate here mainly on comparing the effects of 20-h and 10-h days on the �Landsberg� erecta ecotype. Under short days, more rosette leaves were formed than under long days because the length of the leaf initiation phase was lengthened. However, when comparing leaves at the same position, the leaves on the long-day plants had a greater leaf area than those on the short-day plants. This resulted from greater absolute and relative leaf expansion rates. The number of epidermal cells was also significantly greater under long days but the effect of this was partly negated by the smaller area of the epidermal cells. Thus, the conditions that induce or inhibit flowering also affect leaf development but are the two developmental processes connected? Transferring plants from 10-h to 20-h days induced flowering and inhibited initiation of rosette leaves, consistent with the earlier experiments. To study the effects of flowering delay, the authors grew plants under 14-h days (in which flowering does occur) but then removed the flower buds. The results presented are confined to epidermal cell dynamics: predictably, the epidermal cell area was increased by this treatment but, perhaps unexpectedly, epidermal cell numbers were also increased. Nevertheless, the overall picture is clear: there are whole-plant mechanisms associated with flowering that contribute to the control of leaf development.www.aob.oxfordjournals.org
Effect of water and oxygen:
By just treating the seedling with cold temperature, the said structures do not get verbalized. Along with the cold treatment plants also require water and oxygen for effective vernalization. The seeds or embryos should possess at least 40-50% water in their cells, without which cold treatment has no effect. Similarly oxygen is very essential; probably it is required for biological oxidation. The essentiality of carbohydrates for effective vernalization supports the view of requirement of oxygen. Still, it is difficult to explain how cells use carbohydrates and oxygen for enzymatic oxidative process at such low temperature.
Vernalin:
Various experiments in the past have revealed that during cold treatment, the meristematic cells found either in stem apex or leaves are stimulated to produce some substance. The presence of such substance has been demonstrated by grafting a vernalized plant to another non vernalized plant at normal temperatures. The plant that receives the graft, after sometime, starts producing flowers, which suggests some substance found in the vernalized plants is transported to non-vernalized plant and that is responsible for the induction of flowering in the latter plant. In some cases, if the cut shoot tip of the vernalized plant is placed above the decapitated stem of the non vernalized plant, flowering is induced in the receiver plant The above experiments clearly demonstrate that some substance (X) is synthesized in vernalized plants now called ‘Vernalin” and it is capable of diffusion and induce flowering. Attempts to isolate and identify the components of vernalin have failed. Whether the vernalin is the same as florigin or a precursor of florigin is not known.
Devernalization:
If vernalized seedlings or seeds are subjected to higher temperature like 35-40 0C the plants fail to flower. Such a nullifying effect by higher temperatures is called Devernalization. Nevertheless, if the vernalized plants are maintained at sufficiently low temperatures for a little long period of time, which has to be determined for every species, devernalization is not possible. This may be due to the putative vernalin might have already moved and acted upon the genetic material and committed it in flower formation. However, devernalized plants can be revernalized by subjecting the same seedling or seed again for another period of cold treatment by repetition of vernalization and devernalization cycles. When vernalization is prolonged the seedlings loose the effect of vernalization and lose their viability and potentiality to flower.
The effects of high temperature interruption during low temperature treatment on inflorescence formation in turnip plants (Brassica napa L. cv. Hikari) were investigated. Plants were exposed to 9℃ (low temperature treatment) with interruptions of 17℃, 24℃ or 30℃ for 4, 6 or 8 hr per day. After the treatment, flower formation indices (%) were calculated. Based on these data, the total hours of vernalization, and hours of vernalization or devernalization per day were estimated. Flower formation indices were lower in the group of plants exposed to high temperature interruption than in the group of plants exposed to continuous low temperature. Moreover, the higher the interruptive temperature became and the longer its duration per day, the lower the flower formation index became. The indices were particularly low, when turnip plants were exposed to temperatures of 24℃ or 30℃ for 6 or 8 hr per day. The influence of high temperature interruption during the low temperature treatment was determined quantitatively by measuring the estimated hours of vernalization and devernalization per day. For example, vernalization effect of 9℃ was diminished by 50 %, when the plants were exposed to 17℃ for 8 hr per day during the low temperature treatment or to 24℃ or 30℃ for 4 hr per day during the low temperature treatment. Moreover, the vernalization effect of low temperature treatment was diminished by 90 % when plants were exposed to 24℃ or 30℃ for 8 hr per day during the 9℃ treatment. This can be called as Devernalization.
Gibberellin as substitute for vernalization:
Many plants which require cold treatment also require proper photoperiodic treatment for the induction of flowers, without which vernalization does not have any effect. If such plants are treated with gibberellins (GA), they produce flowers without subjecting the plants to cold and photoperiodic treatments. For example, Henbane is a rosette leaved long day plant which requires cold treatment for flowering. If such untreated plants are sprayed with GAs, the plants produce flowers. It means gibberellins not only substitute vernalization but also photoperiodic treatment. But the same gibberellins have no effect on other long day and cold requiring plants species like Petkus rye. In some cases for the proper response to GA treatment, the plant should possess a cluster of leaves in rosette form as a precondition. The effect of GA on plants like Henbane has been attributed to its effect on the elongation of internodes at which time GA also promotes and probably elaborates the factors required for the induction flowers.
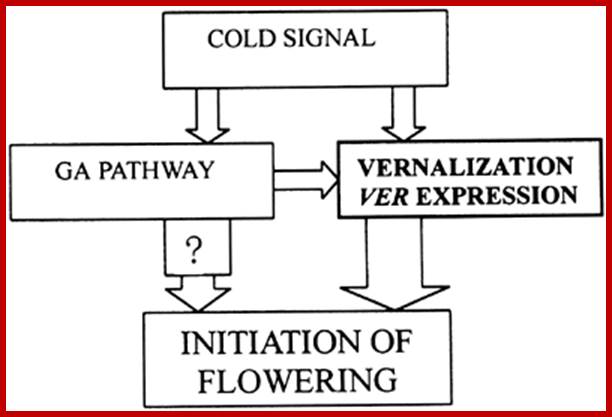
Role of GA in vernalization pathway:
Cold treatment activates VER203 gene expression. GA also induces VER203 expression thus GA can partially substitute cold treatment to initiate flowering in plants that require cold treatment.
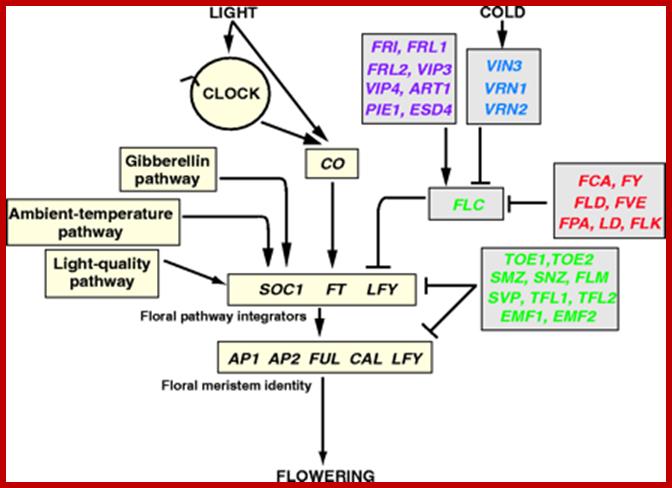
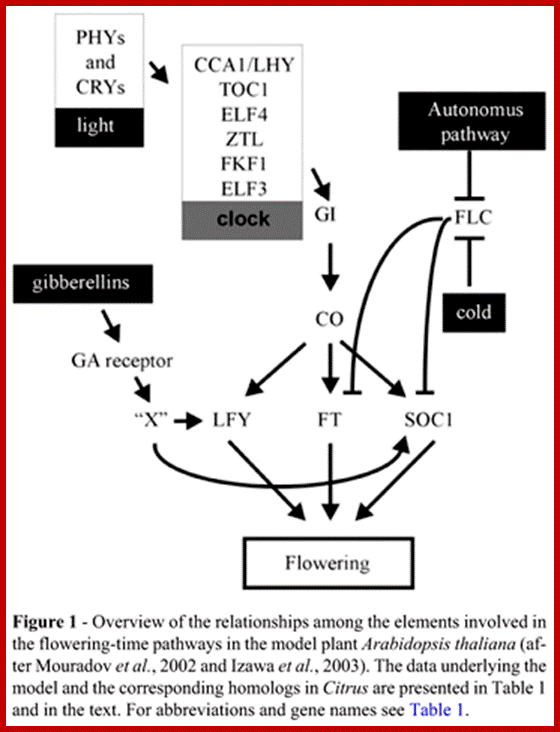
Pathways controlling flowering-time inArabidopsis. The flowering-time pathways control the expression of the floral pathway integratorsSUPPRESSOR OF OVEREXPRESSION OF CONSTANS1(SOC1), FT and LEAFY (LFY). These genes encode proteins that activate the floral meristem identity (FMI) genesAPETALA1 (AP1), APETALA2 (AP2),FRUITFULL (FUL), CAULIFLOWER (CAL) and LFY, which convert the vegetative meristem to a floral fate. Recent expression data has indicated that FUL may also act as a floral integrator (Schmid et al., 2004). The photoperiod, gibberellin, light-quality and ambient-temperature pathways activate floral pathway integrators. The CONSTANS (CO) transcription factor functions in the photoperiod pathway; long-day photoperiods promote flowering by circadian clock (CLOCK) dependent and independent mechanisms, which control the activity of CO. Activation of flowering is antagonized by the floral repressors encoded by (shown in green) FLOWERING LOCUS C (FLC), FLOWERING LOCUS M (FLM), TERMINAL FLOWER1 (TFL1), TERMINAL FLOWER2 (TFL2), SHORT VEGETATIVE PHASE (SVP), TARGET OF EAT1 (TOE1), TARGET OF EAT2(TOE2), SCHNARCHZAPFEN (SNZ), SCHLAFMUTZE (SMZ) and EMBRYONIC FLOWER1/2 (EMF1, EMF2). TFL1 may also be downstream of CO, as it is induced after CO activation (Simon et al., 1996). FLC expression is controlled by a number of different pathways. The genes shown in purple, FRIGIDA (FRI), FRIGIDA-LIKE1(FRL1), FRIGIDA-LIKE2 (FRL2), PHOTOPERIOD INSENSITIVE EARLY FLOWERING1 (PIE1), AERIAL ROSETTE1 (ART1), EARLY UNDER SHORT DAYS4 (ESD4), VERNALIZATION INDEPENDENCE3 (VIP3) and VERNALIZATION INDEPENDENCE4 (VIP4), encode proteins that promote FLC expression and delay flowering. FLC expression is down regulated in response to prolonged cold by proteins encoded by the genes (shown in blue) VERNALIZATION INSENSITIVE3 (VIN3), VERNALIZATION1 (VRN1) and VERNALIZATION2 (VRN2), and also by proteins encoded by the genes of the autonomous pathway (red): FCA, FY, LUMINIDEPENDENS (LD), FLOWERING LOCUS D (FLD), FVE, FLOWERING LOCUS K (FLK) and FPA. The distinction between potential transcriptional and post-transcriptional functions of genes of the autonomous pathway is not made here, but is shown more clearly in Fig. Ian R. Henderson and Caroline Dean; http://dev.biologists.org/
Mechanism of floral induction in vernalized plants:
It is clear from the earlier discussions that the plants with their specific genetic makeup respond to different treatments like cold or photoperiods and produce flowers. Most of the cold requiring plants also require proper photoperiodic treatment. Gibberellins are known to overcome both cold treatment and photoperiodic treatment in long day plants, but it has no effect on short day plants. Synthesis of some unknown substance called ‘Vernalin’ during the period vernalization has been clearly demonstrated by grafting experiments. Furthermore for proper vernalization, plants require sufficient amount of water, oxygen and some vegetation growth. Though all the above said factors are provided to the plant, flower inducing substance won’t be synthesized until and unless it is treated with proper cold condition at the stage of its development. It is during the cold treatment, the synthesis of the said flowering inducing factor is believed to be accelerated.

The GA-deficient na-1 mutant elongates dramatically when grafted to a NA stock with mature leaves of pea plant. The na-1 plant on the left was ungrafted, and remained short. www.textarchive.ru


Arabidopsis; Winter annual requires cold treatment for flowering but Rapid recycler does not require vernalization; Plant Physiol; http://www2.warwick.ac.uk/
According to the hypothesis proposed by Purvis et.al (older view), plants normally synthesize a substance called ‘B’ from some unknown precursor called ‘A’. The synthesis of B is accelerated by cold treatment; when ‘B’ is synthesized; it persists for a period of time. Further conversion of B to C and then D is under the control of photoperiods. The substance D is supposed to be the flowering hormone and C an intermediate compound. The conversion of B to C is slow, but once it is formed it is rapidly converted to D. Once D accumulates to a critical level, it induces flowering. If the same vernalized plant is kept under short day conditions the conversion of B to C is inhibited, instead C is converted to B and then B is converted to E. So the plant remains in vegetative conditions. In fact today scientists have shown that cold treatment induces the expression of Constans (CO) which in turn induces a substance called flowering locus T (FT), which with FD induce the expression of floral meristem identity genes. These on expression act on floral part inducing genes.

The continuity and plasticity of plant development rely on the regulation of meristem activity in response to endogenous and environmental signals. Many plant development regulators involved in meristem function are transcription factors or signalling molecules. In the past few years, the role of chromatin remodeling in programming, maintaining or resetting specific gene expression profiles in subsequent cell generations has been shown to be crucial in plant development. Here, we summarize plant chromatin-remodeling factors required to regulate shoot apical meristem activity, particularly its maintenance during organogenesis and transitions between distinct developmental phases. www.cell.com
This particular hypothesis is based on the studies on ‘Petkus rye’. This plant even if it is not vernalized and grown under normal temperature and if proper photoperiodic treatment is given, it still flowers, but it takes a long time. On the contrary, if it is subjected to vernalization and then subjected to long photoperiod, it produces flowers in a shorter time. It means that the synthesis of A to B takes place all the time, but it is accelerated under cold treatment. Once B accumulates in sufficient amounts, B is converted to C and then to D, which is actually under the control of long day conditions. But in short day or day neutral conditions C is converted back to B and then to E which keeps the plant in vegetative conditions. http://www.scielo.br/
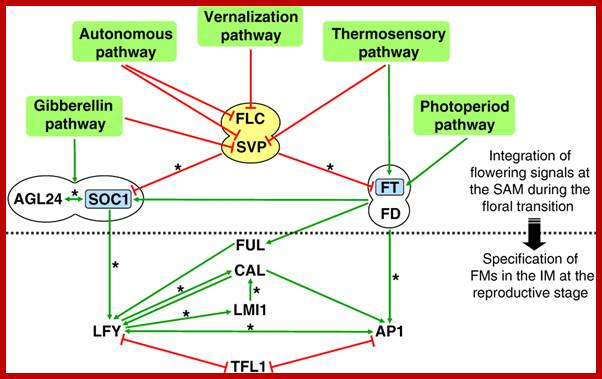
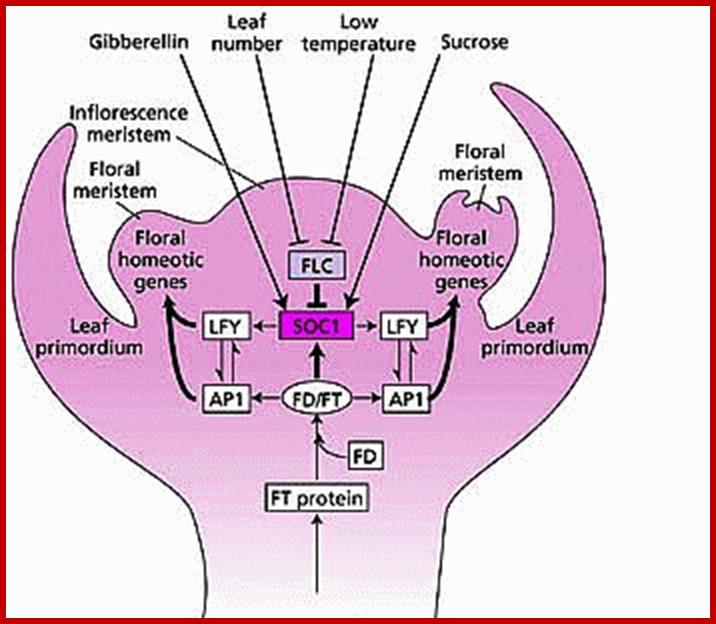
Coming to Bloom; specification of floral meristems. Regulation of FM identity. FM identity is regulated through the integration of multiple flowering signals, with the floral pathway integrators SOC1 and FT (blue) perceiving environmental and developmental signals through several flowering genetic pathways. During the floral transition, the FLC-SVP complex (yellow) repressesSOC1 expression in the leaf and SAM and FT expression in the leaf, whereas the FT-FD complex promotes the expression of SOC1, AP1, and probably FUL in the SAM. SOC1 and AGL24 directly upregulate the expression of one another and also form a protein complex, which is localized at the SAM. In the IM, the increased activity of SOC1 and FT promotes the expression of several FM identity genes, includingLFY, AP1, CAL and FUL, which in turn specify FM identity on the flanks of the IM. Green arrows indicate promoting effects, whereas red linkers indicate repressive effects. Two linked ellipses indicate protein-protein interactions. Asterisks indicate direct transcriptional regulation. AGL24, AGAMOUS-LIKE 24; AP1, APETALA1; CAL, CAULIFLOWER; FLC, FLOWERING LOCUS C; FM, floral meristem; FT, FLOWERING LOCUS T; FUL, FRUITFULL; IM, inflorescence meristem;LFY, LEAFY; SAM, shoot apical meristem; SOC1, SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1; SVP, SHORT VEGETATIVE phase ;http://dev.biologists.org/
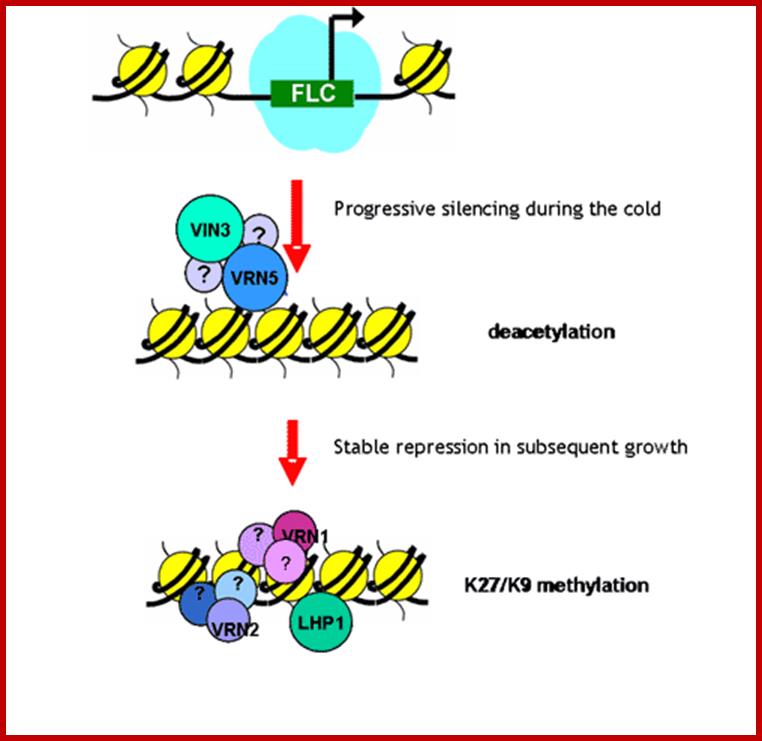
FLC represses genes for flowering by histone tail deacetylation and methylation and after cold treatment demethylation and acetylation leads to activate flowering genes;www.pcb.org
Though this hypothesis appears to explain some observed phenomenon in petkus rye a long day plant, it does not offer any explanation how short day plants flower what is the nature of the intermediate compounds. There are more ambiguities in this theory, than it explains.
Chailkhyan, on the other hand is of the view that floral induction requires two substances. One is GA or GA like compound and the other is flower inducing substance. So he assumed that cold requiring long day plants contain enough flowering substance but lack in GA like compound which will be synthesized under inductive conditions. Contrary to this, cold requiring short day plants possess sufficient amount of GA like compounds and lack in flowering substances which will be synthesized during inductive conditions. The GA like compound and flowering substance combine to produce a flower inducing complex called florigin. Whether it is a cold requiring plant or photoperiod requiring plant or a plant which requires both, the hormone that is required for flowering has to be the same. Chailkhyan’s view was supported by G. Melcher, who grafted a short day Maryland mammoth plant (it does not require cold treatment), to another non–vernalized cold requiring long day Hyoscyamus plant. Grafting resulted in the formation of flowers in Hyoscyamus plant even under non inductive conditions.
. 
http://pixgood.com/
The explanation is that each of these plants possesses one of the two essential substances and lacked the other, i.e., short day plant contains GA and the long day plant possesses the flowering substance. Due to grafting the GA from short day plant diffuses into long day plant, where both combine to produce a flower inducing substance or florigin, which inturn induces. In spite of numerous efforts to isolate and identify the vernalin or the flowering hormone, have remained elusive. Presence of some such substances has been established by experiments but the nature of that material is not known. More than that, nothing is known about the molecular events that lead to flower formation in response to cold treatment. Whether cold treatment induces any gene expression or whether cold treatment destroys some repressor proteins that are bound to MRNAs there by activities pre-existing MRNPs, whether vernalization brings about the destruction of a repressor, there by activities a gene expression are few of the questions for which there is no answer. But one thing is certain that during vernalization period some thermo labile component is produced and it remains for a long period of time in the plant and it works with another substance produced during photoperiodic conditions.
Actually plants requiring vernalization contain FLC protein bound to specific location in chromatin that represses genes required for flowering, and it keeps in repressed state till cold treatment. It is during cold treatment the FLC dissociates from the loci and allow other factors to bind and activate gene expression. It is this that makes the plants to flower in response to vernalization.

Model for the regulation of the floral repressor FLC throughout the Arabidopsis life cycle; During seedling growth, a group of genes encode proteins that function as activators of FLC expression (shown in purple); these genes include FRI, FRL1, FRL2, ESD4, ART1, PIE1, VIP3 and VIP4. These proteins may maintain FLC chromatin in an active state (indicated by an open structure and the presence of active histone tail modifications shown in green). The autonomous pathway functions antagonistically to the activators to repress FLC expression. The RNA-binding proteins FCA, FPA and FLK, and the polyadenylation factor FY, may function post-transcriptionally to achieve this and are shown in red. The FVE/FLD proteins act with a putative histone deacetylase (HDAC; all shown in orange) to promote an inactive FLC chromatin state, represented by a closed structure with inactive histone tail modifications (red). FLC is also repressed by exposure to long periods of cold (vernalization). The proteins acting in the vernalization pathway are shown in pink. Prolonged cold induces VIN3expression, which promotes an inactive FLC chromatin state. Subsequently, the VRN1 and VRN2 proteins are recruited to FLC, and are required for the methylation of FLC histones and the maintenance of silencing. These marks may promote the association of silencing factors with FLC chromatin that reinforce its repression. During meiosis, gametogenesis or early embryogenesis, FLC repression is overcome, thus resetting its expression in the next generation. Ian R. Henderson and Caroline Dean*dev.biologists.org
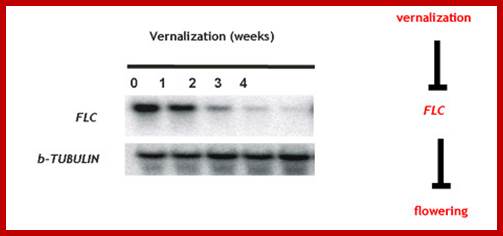
During cold treatment, with time the concentration of FLC, the repressor goes down, means its binding to chromatin gets reduced with time under cold treatment.www.pcb.org
Arabidopsis as a model system is used to study the regulation of flowering time. In addition to the general advantages (small sized, rapid generation time, sequenced genome, and a large collection of gene knock-out lines), flowering in Arabidopsis is regulated by both photoperiod (flowering is accelerated by long days) and vernalization. Early-flowering lab strains of Arabidopsis do not have a vernalization requirement for early flowering; however, naturally occurring winter-annual accessions of Arabidopsis are late flowering and vernalization responsive due to dominant alleles of the FRIGIDA (FRI) gene. A number of late-flowering vernalization-responsive mutants (such as FLD) have also been isolated from mutagenized early flowering strains. The genes identified by these mutants are collective known as the autonomous pathway floral promotion pathway.
FLOWERING LOCUS C (FLC) blocks flowering prior to vernalization;
Work has shown that elevated levels of the MADS-domain-containing transcription factor FLC cause the late-flowering vernalization-responsive phenotype of naturally occurring strains containing FRI or autonomous pathway mutants. FLC acts to block flowering and FRI, in turn, acts to up regulate FLC levels. The autonomous-pathway genes promote flowering by repressing FLC expression; thus autonomous pathway mutants contain elevated levels of FLC and are late flowering.
Vernalization promotes flowering by causing an epigenetic shut off of FLC:
Our work has also demonstrated that the promotion of flowering by vernalization is also regulated by FLC. The late flowering phenotype of FRI or autonomous pathway mutants can be effectively suppressed by vernalization. This suppression takes place at the level of FLC regulation. Following vernalization FLC expression is down regulated; for the rest of the plant's life and FLC levels remain suppressed. Levels are then reset during reproduction and are high again in the next generation.
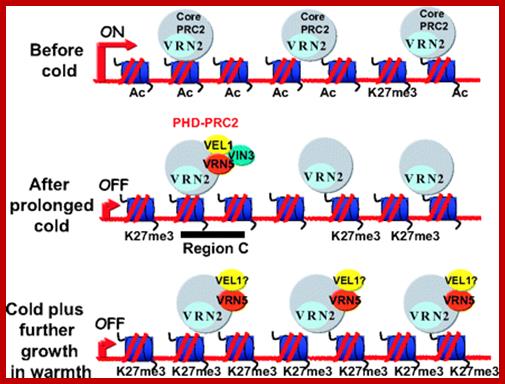
Silencing or reducing the concentration of VRN during cold treatment is shown, where VEL1, 2 and 3 bind to VRN at C-locus to offset effect of chromatin silencing in the locus. Possible sequence of events in the assembly and association of specific Polycomb complexes at different stages of epigenetic silencing of FLC during vernalization is shown. In nonvernalized plants the PRC2 core complex is associated with the whole FLC locus. Prolonged cold leads to VRN5 localization specifically at region C as part of the PHD-PRC2 complex with VIN3 and VEL1 and to a decrease in histone acetylation. In plants returned to warm conditions, VIN3 is no longer expressed, and VRN5 associates more widely throughout the locus, concomitant with a significant increase in H3K27 trimethylation.www.pnas.org
We, authors of the above figure, are currently working to understand in greater detail how FLC is regulated by FRI, the autonomous pathway, and vernalization. We used a variety of genetic and molecular approaches in this pursuit. Recent work by our lab and others has shown that changes in chromatin structure are critical in FLC regulation. One gene recently cloned by our lab, EFS, is required for high levels of FLC expression; efs mutants suppress the late-flowering effects of FRI and autonomous-pathway mutants. EFS encodes a SET-domain containing transcription factor. SET domain proteins are known to act as histone methyl transferases, which can modify chromatin structure and thereby regulate gene expression.
Epigenetics refers to heritable changes in patterns of gene expression that occur without alterations in DNA sequence. The epigenetic mechanisms involve covalent modifications of DNA and histones, which affect transcriptional activity of chromatin. Since chromatin states can be propagated through mitotic and meiotic divisions, epigenetic mechanisms are thought to provide heritable ‘cellular memory’. Here, we review selected examples of epigenetic memory in plants and briefly discuss underlying mechanisms. Mayumi Iwasaki, Jerzy Paszkowski.
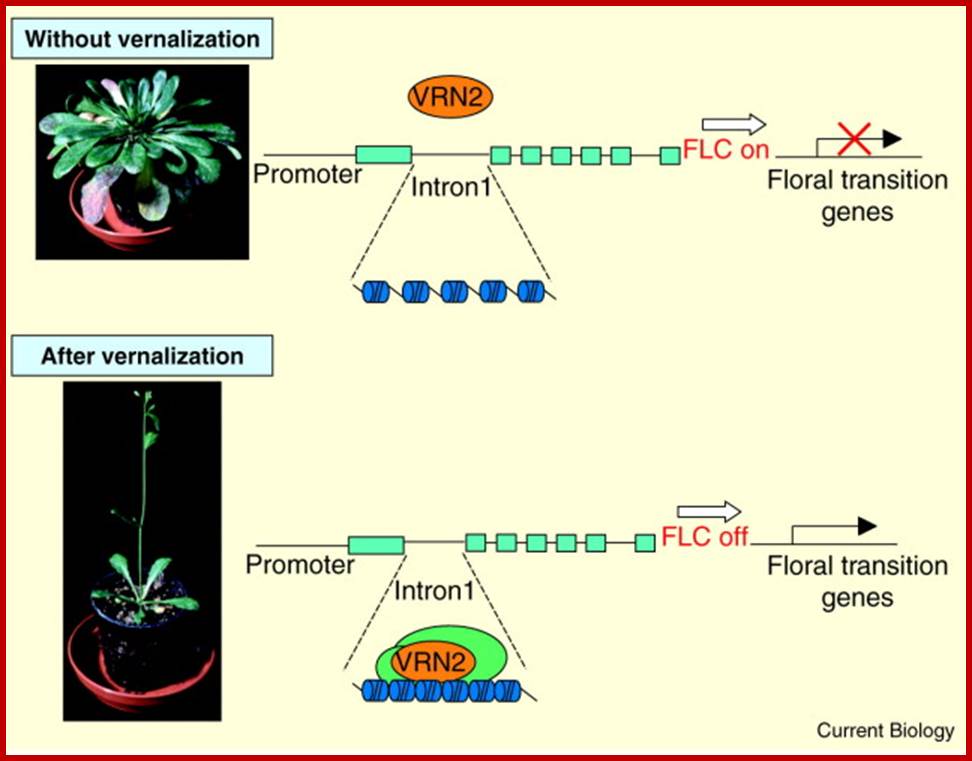
Epigenetics-flower that comes from cold; A model for the action of the Polycomb group protein VRN2 in vernalization, based on the results of Gendall et al. vrn2 mutants show an increased DNAse sensitivity of FLC after vernalization suggesting that VRN2 changes the structure at the FLC locus by recruiting a protein complex with chromatin remodeling activity. This could establish or maintain the epigenetic mark, which enables the plant to remember periods of cold temperature for several weeks. (Pictures courtesy of Caroline Dean.) http://www.sciencedirect.com/
Plants can remember periods of cold several weeks later and respond by switching from vegetative to reproductive development. Recent findings shed light on this phenomenon by showing that a gene responsible for keeping this memory encodes a member of the Polycomb group of proteins.
In flowering plants, the transition from vegetative growth to flowering is controlled by both developmental and environmental signals. Using Arabidopsis thaliana as a model system, four major flowering promoting pathways have been defined genetically. The pathways that respond to photoperiod and ‘vernalization’ — a period of cold treatment that accelerates flowering — promote flowering by integrating environmental signals, whereas the autonomous and gibberellin pathways act largely independent of the environment, but depend on the developmental competence of the plant. Many Arabidopsis ecotypes collected at high latitudes or alpine regions are winter annuals that flower in spring after exposure to winter conditions. In the laboratory, these ecotypes flower very late, but they flower much earlier when the seedlings are exposed to prolonged cold treatment. The requirement for vernalization ensures that flowering occurs in spring, providing the maximal opportunity for seed set.
Vernalization often occurs at the seedling stage, with flowering occurring weeks later. The meristem thus has to remember this stimulus over several cycles of cell division, suggestive of an epigenetic process. It was hypothesized that DNA methylation is the epigenetic mark that enables the cell to recall previous cold conditions. A recent study by Gendall et al has shed new light on the epigenetic basis of vernalization by showing that a Polycomb group protein is involved in the maintenance of the vernalization response above.