ABSORPTION OF WATER
Unlike aquatic plants, terrestrial plants have to absorb water form the soil all the time to maintain turgidity, metabolic activities and growth of the plant. It is essential to understand the structure of soil, its water content and factors; and also the plant structures involved in absorption of water.
Soil structure:
Soil is made up of fine rock particulate of various sizes derived from the weathered igneous and sedimentary rocks. Environmental factors like, heat, wind, rain, cold, river streams and oceanic waves act upon the rock particles, which break them down to smaller particles which on accumulation in a shallow or flat surfaces, constitute the soil. The process of soil formation on this planet is a continuous process and it is taking place for the past 4.0 billion years and will continue as long as this planet exists. The same process also operated on other planets.
Based on the size, structures of soil particles and the composition of organic and inorganic components, soils have been classified into rocky, coarse, sandy clay and loamy soils. Rock particles of big sizes do not hold any water between them and any such soil consisting of rock particles which do not hold water in between them is not good for the development of root system. Even the sandy soil of such small sized rocks is good for aeration but not for water retention. On the other hand, clay soils have colloidal particles that can hold water but very poor in aeration. However, the loam soil is good, because it has the mixture of clay, sand and decomposed organic material called humus. This soil provides good aeration and proper capillary spaces to hold water. Thus this soil is considered to be the best soil for the luxuriant growth of the root system.
SOIL WATER:
Rain water is the main source of water for most of the land plants. When the rain water falls or well water canal or river water flow on the soil, some water percolates into and moves into inter spaces found between rock particulates.
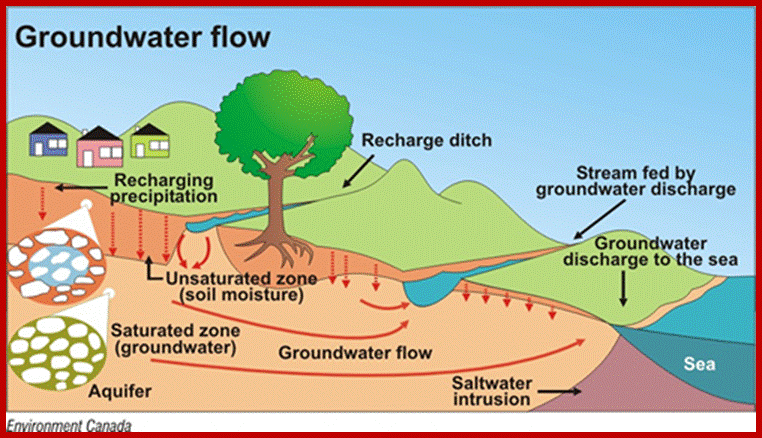
Water finds it gravity and flows in the gradient; www.atsdr.cdc.gov
Water in the soil; www.growflow.com.au
At the same time a lot of water moves all along the slope of the landscape; this water is often called runaway water, which is of no use to the root system. On the other hand, the rain water or any other water that enters into the soil moves downwards. On its way, it fills up all the capillary spaces found in soil and still moves downwards (by gravitational forces) till it reaches the water table. Such water that goes downhill in the soil is called gravitational water. Again, this water is not useful to the root system.
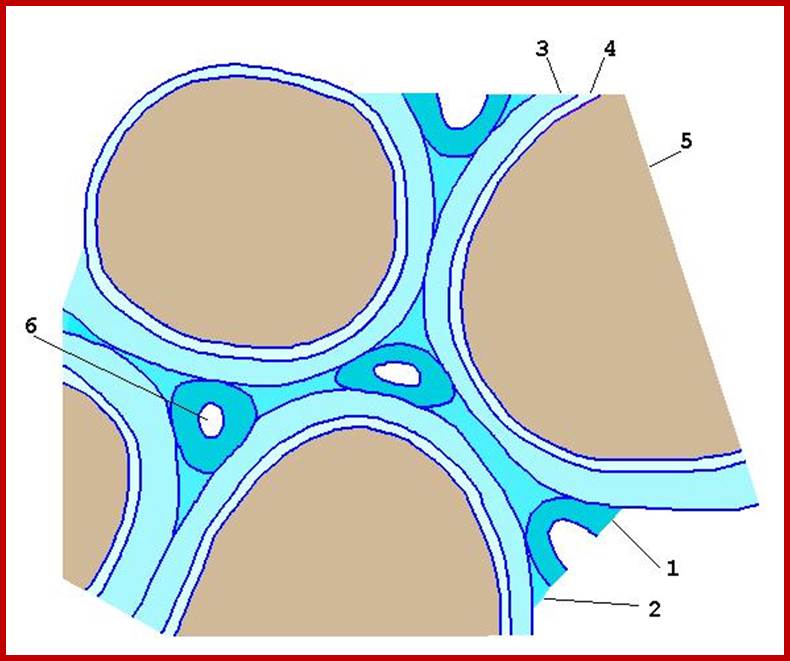
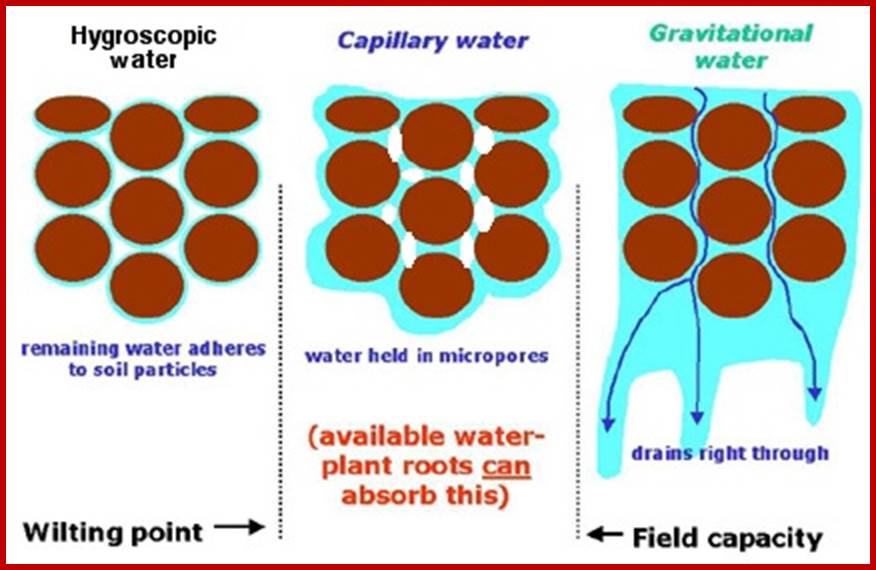
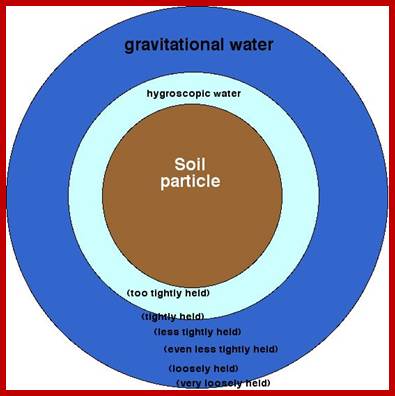
Capillary water 2 in the diagram; the moisture around the soil particles 1,3,4,5 and 6, available for plants and it also contains soluble salts and minerals. http://water-salt.narod.ru/site_en
Each soil particle holds a thin film of water around its surface, it is called hygroscopic water. This water is not available to roots, because this water is tenaciously holding on to the rock surface or soil particle surface. In spite of it, the water that is found in between particles that is the capillary spaces, called capillary water; it is this water that is available to the roots and useful. So, the capillary water is the main source of useful water in the soil for the plants. Moreover, capillary system provides a network of spaces for aeration as well as movement of water from one region to another region within the soil.
FIELD CAPACITY OF THE SOIL
The amount of water found within the soil as capillary water that is available for the root system is often referred to as useful water content of the soil. Hence the capacity of soil to hold maximum amount of utilizable or useful water is known as ‘Field capacity of the soil’, this again depends upon the nature of soil. Sandy and rocky soils are poor soils in terms of water holding potential. While clay soils have great water retention capacity but they have the worst aeration. However, loam soil is the best for it retains good amount of water and also it has good aeration. However, the field capacity of the soil can be determined by finding out the difference between the weight of completely wet soil and that of the dry soil of a known quantity.
WILTING POINT
As roots with their numerous branches and millions of root hairs deplete water from capillary spaces and water from the other regions move into the depleted spaces; sometimes the water moves upwards from the water table and fills up the capillary spaces, but the refilling or replenishment process takes its own time. Thus, plants experience deficiency of water for a short duration time, which is referred to a temporary wilting point. In some cases, the depleted water is not replaced for a long time, under such conditions, plants die, and such a state is called permanent wilting point.
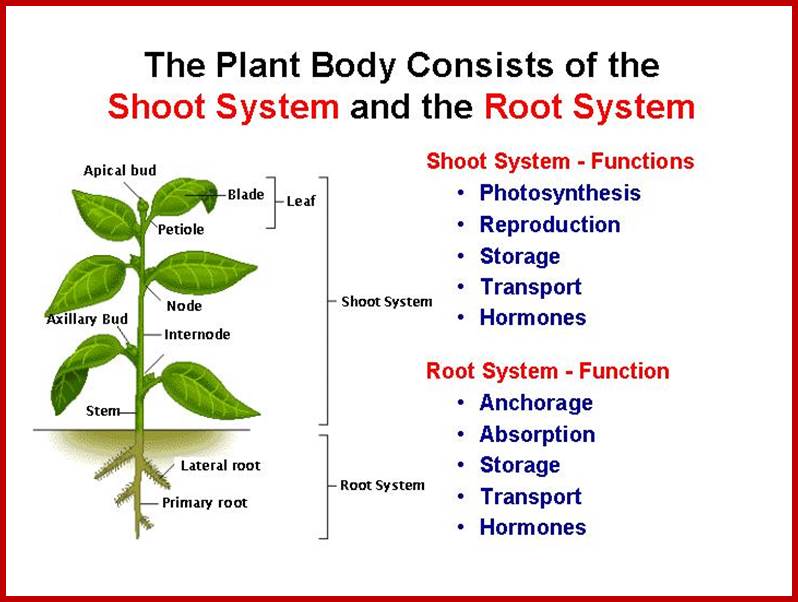
ROOT SYSTEM
Plants growing in aquatic habitat do not require any special structures for absorbing water, but plants growing in soil or growing as epiphytes, posses’ special structures for absorbing water. For example, epiphytic orchids contain velamen root specially adopted to absorb water directly from the atmosphere. But the terrestrial plants produce roots that grow into the soil.

www.svtmontacourbevoie.wordpress.com
Such roots not only fix the plant firmly in the soil but also help in absorbing water and mineral salts from the soil. Besides the above said functions; in different plants, roots also play other functions such as storage, climbing, pneumatophores, etc.
EXTENSIVENES OF THE ROOT SYSTEM
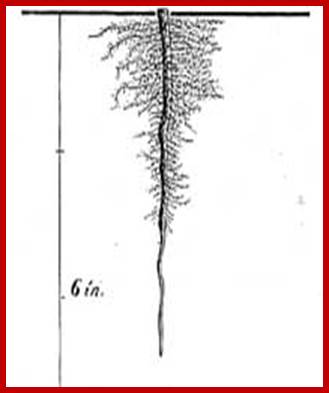
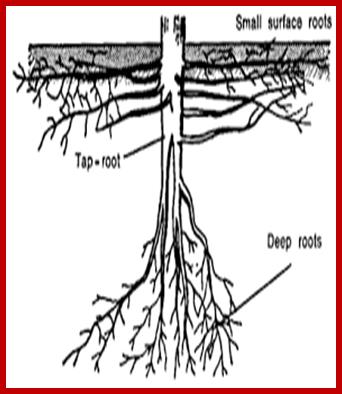
Roots, irrespective of their origin or kind, grow, branch and spread in the soil to a great extent. Some plants produce roots which grow deeper into the soil and reach the water table in some cases.
http://www.fao.org/docrep; cocoa- http://www.fao.org/


http://foodgrowsontrees.blogspot.in/http://soilandhealth.org/
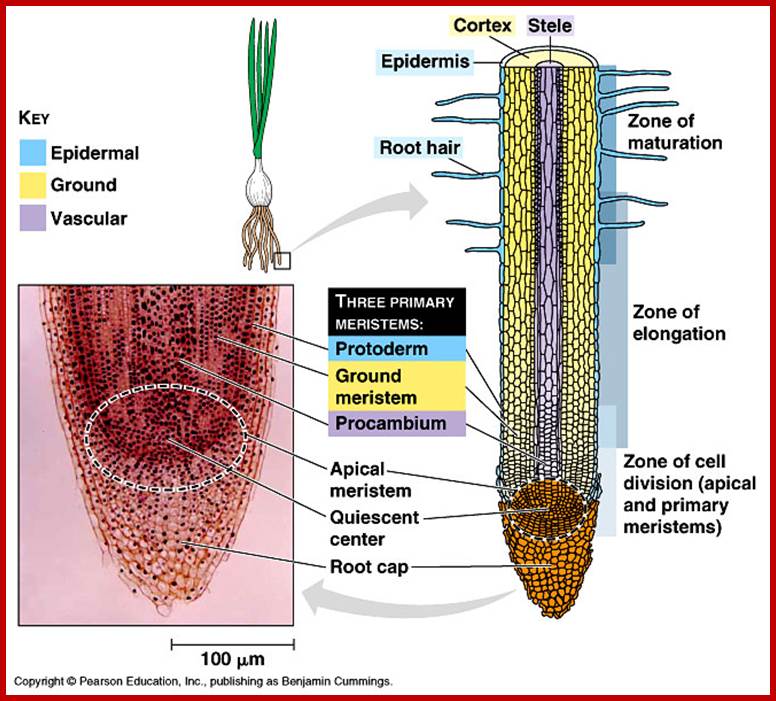
Tap root: Region of root cap; Region of cell elongation; Region of root hairs; and region of maturation; Root Anatomy from Meristem to well-developed vasculature. http://www.askiitians.com/
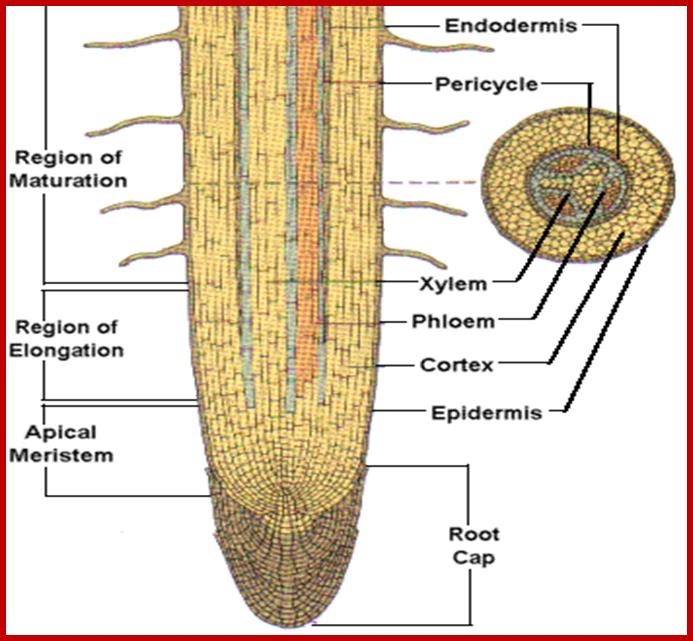
STRUCTURE OF ROOT ENDS.
The terminal region of every root and its branches consists of a region of meristem, region of elongation and region of differentiation. The region meristem is made up of actively dividing cells which by continuous cell divisions produce derivatives. The region of elongation is made up cells derived from the region of meristems and they are in the process of rapid elongation. Next to this, the region of differentiation or the region of root hairs is found. In this region, cells are in the process of differentiation, where some cells are developing into vascular tissues, certain cells specialize into cortical or medullary cells. But a large number of epidermal cells in this region undergo transformation into long tubular structures called root hairs. Beyond the root hair zone, i.e. towards the base of the root, the root exhibits dead root hairs, cortical cells with suberization and most of the cells are fully mature and some of them are even dead. Among the above-mentioned regions root hair zone is mainly responsible for the absorption of water.
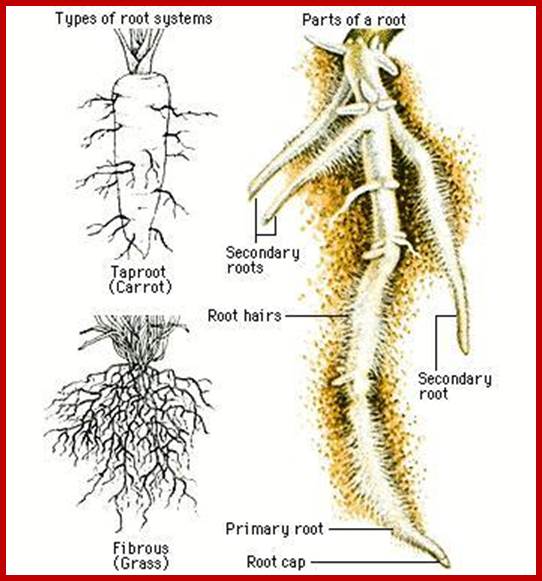
Such roots are called deep feeders. Most of them are tap root systems. But some plants produce branches which spread outwards and do not grow very deep and such roots are mostly adventitious or fibrous roots, they are called surface feeders. However, some fibrous roots also reach great depths.
The development of root system in plants is very extensive in the sense the total length of all the branches put together is amazingly great. For example, a four-month-old corn plant possess an adventitious root system, which if all its branches are put together it extends to about many kms? Trees with their perennial habitats posses’ deep feeder root system and their total length of the entire root system will be more than 200 km? Added to this feature, roots continue to grow every minute and the extent of growth of root. System is in the range of 4-8 kilometers per day?. The ramification of the root system and rapid growth of thousands of root tips is very essential, for the root tip are eternally seeking new areas for the absorption of water and minerals. In spite of its extensive branches and length, the actual region used for absorption of water is restricted to the terminal region of the roots.
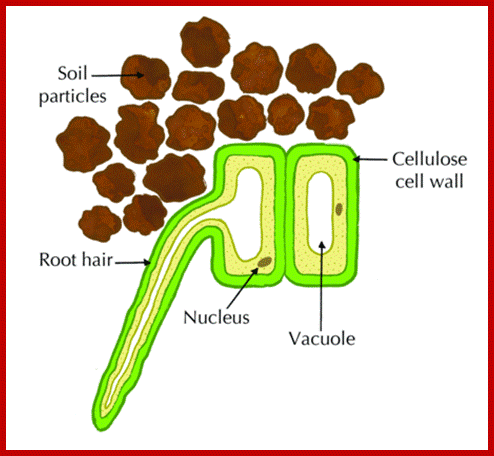
ROOT HAIRS
They are the tubular extensions of epidermal cells found in the region of differentiation, behind the root tips. Each of these root hairs is 0.75 – 1.0 cm in length and 10 μ in diameter.

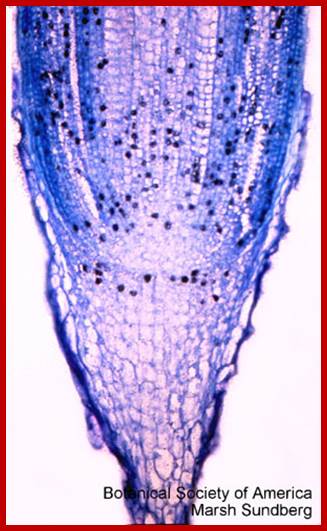
Note relative positions of three primary meristem at the apex; Quiescent center –labeled radioactive nuclei; http://www.bio.miami.edu/
Quiescent center; region of cell division; region of cell elongation and region-maturation; http://www.bio.miami.edu/
http://khodavaelm.blogfa.com/
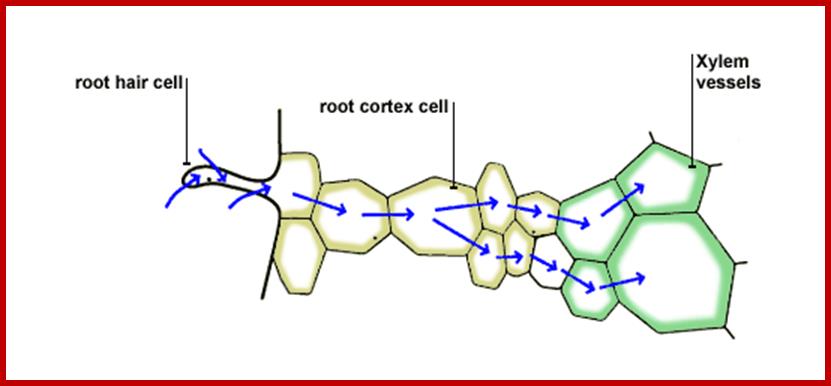
Flow water from soil along root hairs into main roots; www.bbc.co.uk
www.imgarcade.com
http://www.wonderwhizkids.com (for students); ;www.imgarcade.com
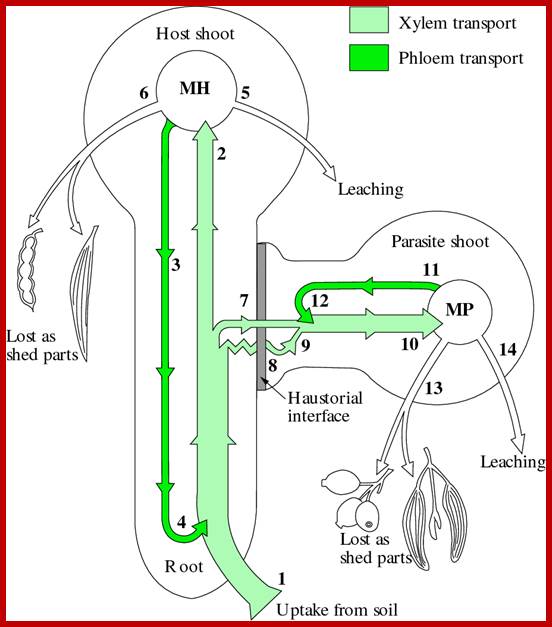
Representation of uptake, transport and partitioning of the phloem-mobile and xylem-tapping nutrient potassium (K+) between a tree host and a xylem-tapping mistletoe. K taken up from soil (1) moves to the host shoot (2) and across the lumen-to-lumen xylem continuities (7) and parenchyma (8) of the hasutorial interface and thence into the mistletoe (10). K in the host shoot is then pictured as being retained in host biomass (MH), lost through leaching by rain (5), in organ shedding (5) or cycled back to the root via phloem (4). Similar accumulation (MP) and cycling (items 11-14) are depicted for the mistletoe but with no backflow to the host. The mistletoe runs at an 'excess' of K due to a higher rate of transpiration and markedly active absorption of K by parenchyma of the hasutorial interface. Foliage shed by mistletoes is still fully loaded with K. (Based on Pate 1995); www.plantsinaction.science.uq.edu.au
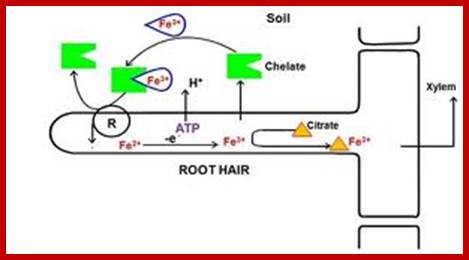
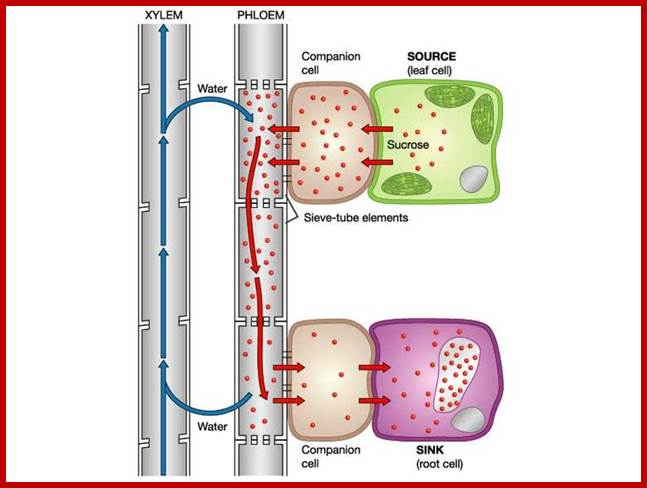
Uptake of Water and Minerals b the roots; m.everythingscience.co.za764
The total number of root hairs present in a four-month-old corn plant is approximately 14 billion or more. Taking the average length and breadth of a single root hair, the total surface area of all the root hairs put together and computed; and it turns out that this surface area absorbs about 28-50 liters of water per day, which is surprisingly 27 times the total amount of water lost from the transpiring surface area. Furthermore, the daily growth and extent of the growth of root hairs, is more than adequate for the amount of water available in the soil. By chance, if there is no replenishment of water that is lost from the soil, the entire volume of soil water can be lost to the atmosphere in about 24-48 hours. This is done through the combined action of total water absorbing surface i.e. roots and root hairs, and transpiring surface area i.e. stomata. If the lost water is not replenished in the soil, plants experience water stress, and if this continues, plants, die, when they reach beyond permanent wilting point. Though more than 90% of the water is absorbed by the root hairs zone, the other regions also absorb the water but the amount of water absorbed by these regions is not much.
Water taken up by the roots of a plant is transported through a plant to the leaves and lost into the air. The stages of the process are:
- Water enters root hair cells by osmosis and also by auaporins. 2. The root hair cell is hypertonic to the surrounding soil water. This means that it has a lower water concentration. 3. Water then moves apoplastically and also symplastically cell to cell through the root cortex by osmosis along a concentration gradient; this means that each cell is hypertonic to the one before it. 4. In the centre of the root the water enters the xylem vessels. 5. Water may move by diffusion through the cell walls and intercellular spaces.
FACTORS THAT CONTROL ABSORPTION OF WATER
Living root system is very essential for the survival of plants, for it is involved in the absorption of water and minerals which is a must. The factors that regulate the health of the roots also control the process of absorption of water. But the most important of them are soil temperature, soil water, soil aeration and the root structure.
ROOT SYSTEM
The efficiency of absorption of water depends upon the extent of root branching and the total surface area of the absorptive structures of root hairs. Though most of the water is absorbed by the root hair zone, other regions also contribute in absorption to some extent. Still the efficiency of absorption varies from one root system to the other.
SOIL TEMPERATURE
In most of the cases, the soil temperature is little lower than that found in aerial regions. As temperature influences the viscosity mobility of water and also the metabolic activity of the plant cells, it affects the ability and the efficiency of absorption of water. The rate of absorption of water is lower, if the temperature is lowered this is because the mobility of water is decreases and the viscosity of liquid water increases. Thus low temperature resists the free movement of water which in turn affects the rate of absorption. However a high temperature has adverse effect on the root’s efficiency.
SOIL WATER
Soil water is not pure water but it consists of a large number of minerals ad organic compounds in dissolved state, so it is a solution. The osmotic concentration of soil solution under full field capacity is always many folds lower the osmotic concentration of cell sap. This provides a kind of osmotic gradient between the soil solution and root cells. When conditions are favorable for rapid transpiration, and shortage of water in the soil, plants exhibit wilting features.
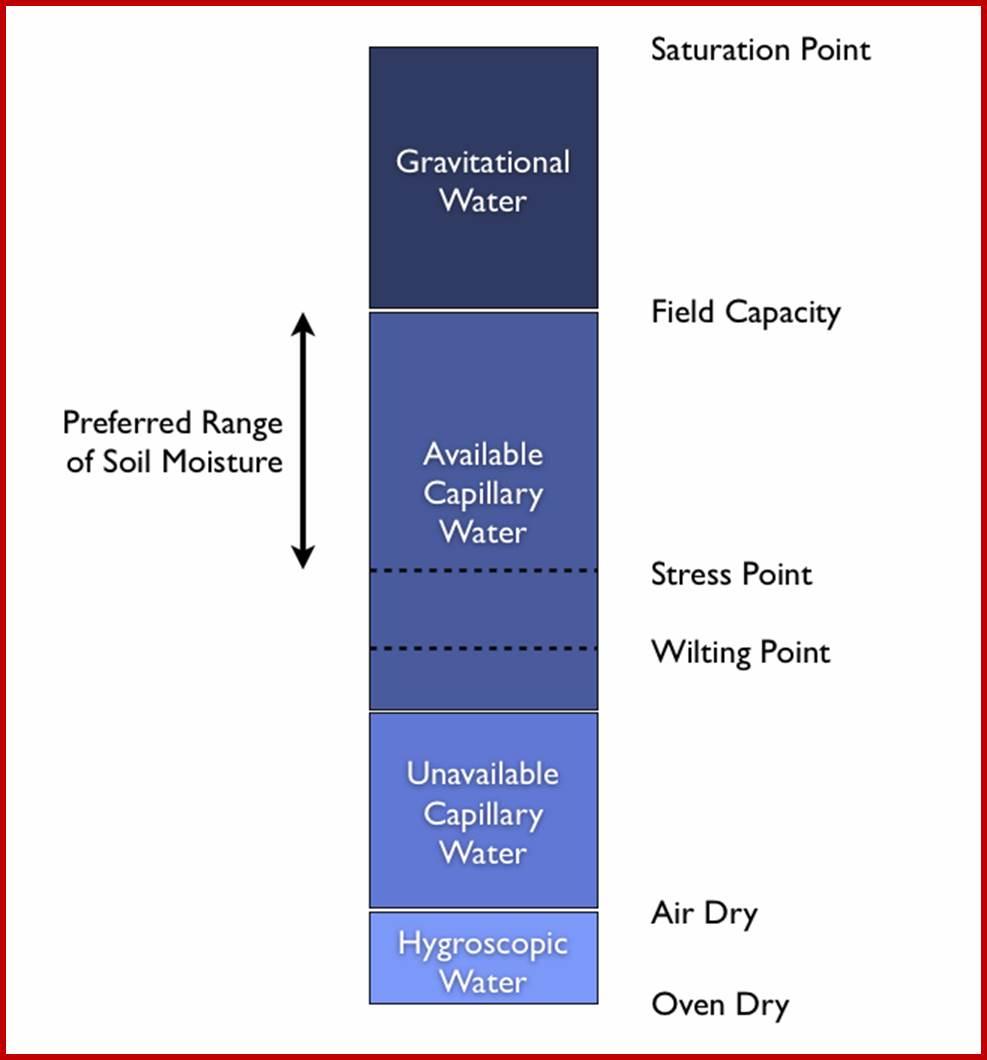
Types of water in the soil; http://redgardens.org/
If the water is not replaced within a particular period of time, plants experience permanent wilting stress and they may die. In fact, under normal conditions, the rate of absorption of water shows diurnal rhythm i.e., higher rate of absorption during day and low rate at nights. Thus, soil water and its constituents determine not only the rate of absorption but also the amount of water absorbed.
AERATION OF SOIL:
Soil being made up of fine rock particles of different dimensions, clay particles and other components, possess plenty of lung space within which air is present. If the soil is water logged, most of the air is expelled from the capillary spaces and roots experience anaerobic conditions and their metabolism suffers. This affects the growth of the roots. Sometimes, excess of clay particles also clog the spaces and soil is rendered unsuitable for the normal growth of the root. Some plants are adapted to grow in water logged areas. In such situations the rate of absorption of water and mineral salts is greatly affected. Even greater accumulation of CO2 within the soil causes change in the pH of the soil solution. Such changes will be detrimental to the root system. But normally the soil CO2 is replaced by the atmospheric air.
PATHWAY OF WATER IN THE ROOT SYSTEM:
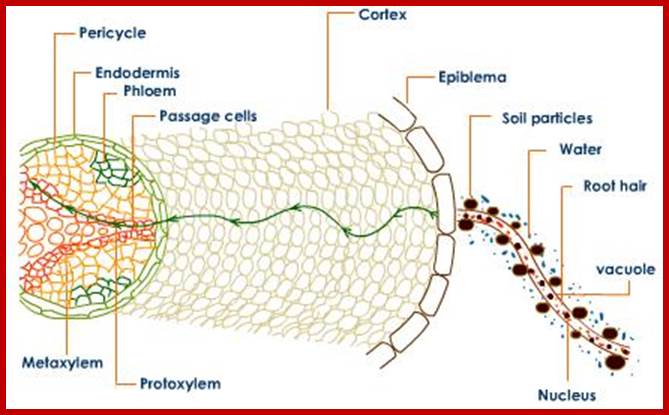
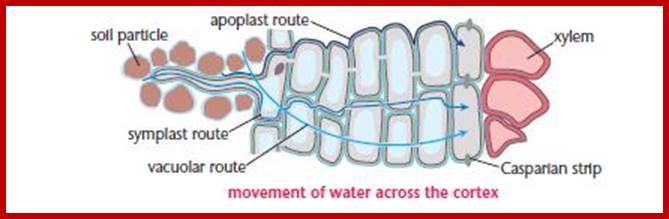
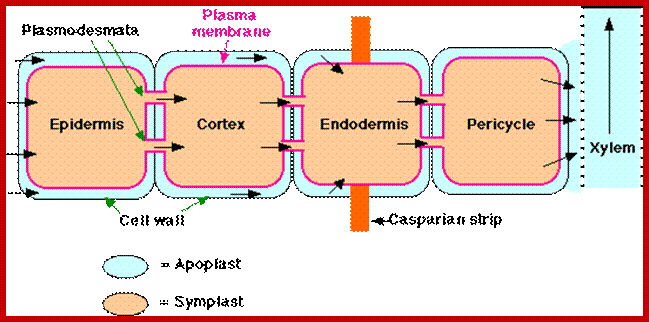
Root is a highly organized multicultural structure containing a variety of cells, some are living and some are dead. Cell wall materials like cellulose, hemicellulose, lignin, etc., form the bulk of inert organic materials of the root cells. In between the micro fibrils and macro fibril part of the cell wall plenty of fine free spaces are found through which water and mineral salts can easily diffuse in and diffuse out. Similarly, the intercellular spaces constitute 6-10% of the total volume of the root system. These spaces also act as spaces for free diffusion of water and other components. Such spaces which are free for diffusion of water and salts are called Apparent Free Spaces (AFS). More over, the network of such spaces provides a continuum from the outer surface of the roots up to the central vascular cylinder. Depending upon the water potential or osmotic gradient, if it is favorable, water from the soil easily diffuses into roots through AFS of cell wall and intercellular spaces. Such movement is called Apoplast movement and it is always very rapid.
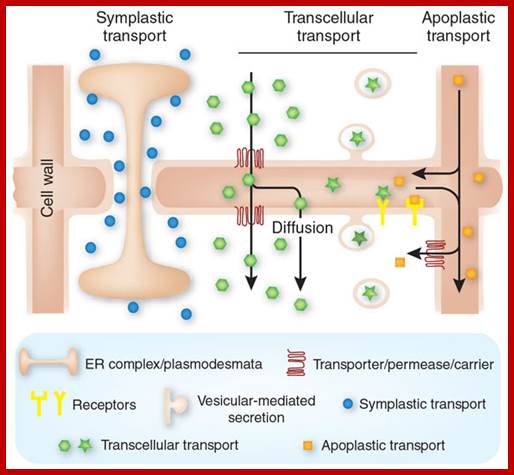
Symplastic transport (left) uses plasmodesmata that interconnect the cytoplasm’s of neighboring cells. A modified endoplasmic reticulum (ER) forms part of the plasmodesmata structure. Apoplastic transport (right) involves passive diffusion of molecules in the extracellular space, the cell wall. Transcellular transport (center) combines apoplastic transport with a secretion- and endocytosis-based or channel- and carrier-based transport pathway to cross plasma membranes.;www.nature.com.
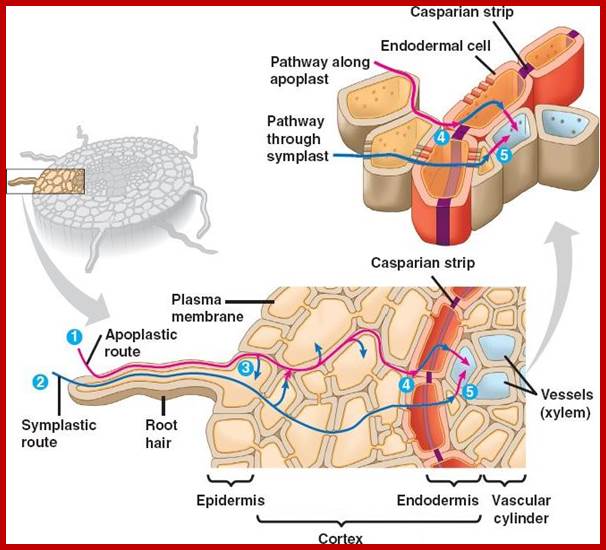
Lateral transport in Roots; bio1903.nicerweb.com.
The area of young roots where most absorption takes place is the root hair zone. The root hairs are delicate structures which get continuously replaced by new ones at an average rate of 100 million per day. The root hairs lack cuticle and provide a large surface area. They are extensions of the epidermal cells. They have sticky walls by which they adhere tightly to soil particles. As the root hairs are extremely thin and large in number, they provide enormous surface area for absorption. They take in water from the intervening spaces mainly by osmosis.
1. Root hairs absorb water and minerals, which move along the walls comprising theapoplast; 2. Minerals and water that cross the plasma membranes of root hairs enter the symplast. 3. Along the apoplasts some water and minerals are transported into the cytosols of cells and then move via the symplast. 4. The Casparian strip, a belt of waxy material, allows only minerals in the symplast to pass into the vascular cylinder through the plasma membrane of endodermal cells. 5. Cells in the vascular cylinder transport water and minerals throughout the plant’s length and breadth.
http://www.slidesshare.net
Communication between the root and the shoot forms a central part of the coordinated response of plants to drought. Recent evidence suggests that a K+ channel expressed in the stelar tissue of the root may have a major role in this process.
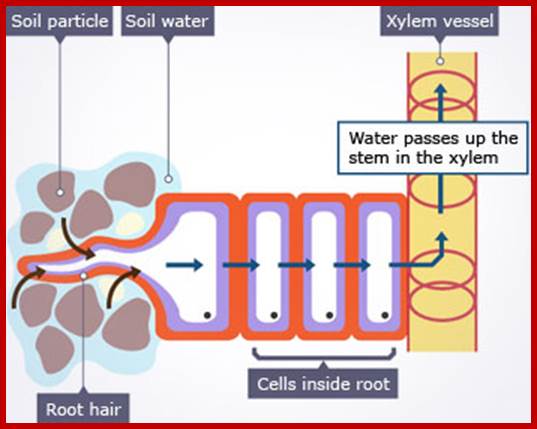
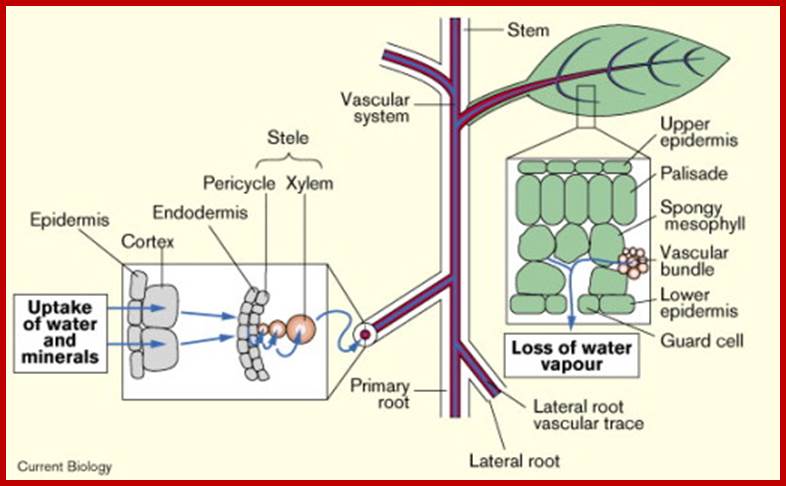
The transpiration stream in a plant; The pathway followed by water and minerals from the soil, through the root into the vascular system and hence to the leaves, is indicated by the blue arrows. The whole process is driven by the loss of water vapors from the stomatal pores on the surfaces of the leaves.
http://www.sciencedirect.com/
Water absorbed, water translocated along the stem and water lost as water vapors.
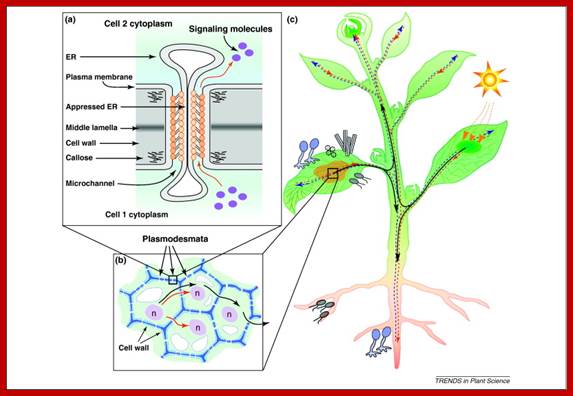
The role of plasmodesmata in the Symplasmic pathway in plants: (a) A schematic diagram of a plasmodesma illustrating the ultrastructure and cell-to-cell trafficking of diffusible signaling molecules. Orange-yellow spheres and short rods represent hypothetical proteinaceous and filamentous components observed within plasmodesma. (b) Plasmodesmal-mediated signaling among symplastically connected cells. Some signals (red arrows) move only into cells adjacent to the original cell that generated them for local communication, whereas systemic signals (black arrows) move farther to reach phloem for long-distance communication. (c) Environmental signals (e.g. day length or light intensity) or challenges (e.g. biotic stresses caused by microbial pathogen infection) perceived in the leaves are processed in the receptive cells (dark-green or yellow patch, respectively) and transported through plasmodesmata for local communication within a tissue. These signals can then enter phloem (broken red arrows) for inter-organ signaling and are transported to distantly located target cells and tissues, such as the shoot or root tips, to bring about appropriate biochemical, physiological, and/or developmental changes. Broken blue arrows indicate xylem transport. WWW.cell.com
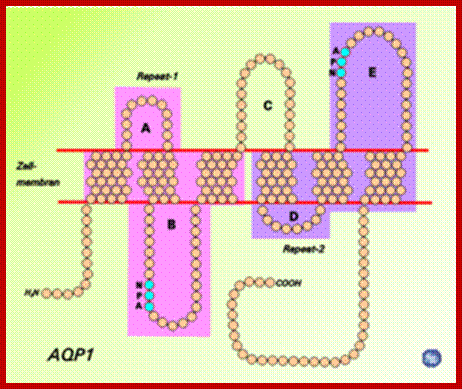
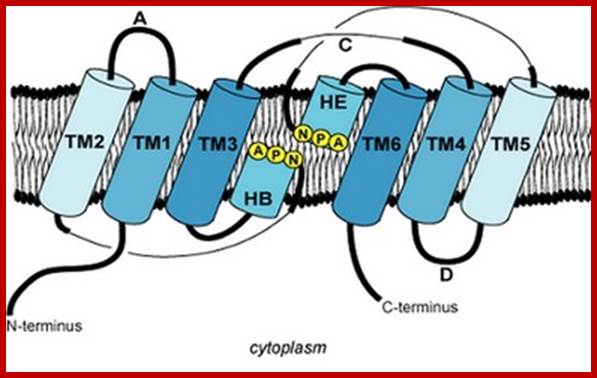
Schematic diagram of the 2D structure of aquaporin 1 (AQP1) depicting the six transmembrane alpha-helices and the five interhelical loop regions A-E. www.chemie-schule.de180 × 151Search by image;
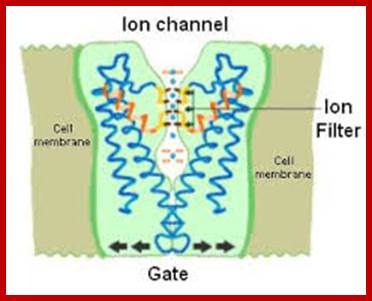
The Kcs-A K+ channel; Fully or partially hydrated potassium ions (blue) are seen just below and above the selectivity filter. In the selectivity filter, potassium ions are coordinated by oxygen atoms (black) in the protein backbone. The channel is closed by a gate in the lower part(arrows). The gate can be opened by sensor domains (not shown) that pull the gate open.;www.nobelprize.org 2003.
The 2003 Nobel Prize in Chemistry was awarded jointly to Peter Agre for the discovery of aquaporins, and Roderick MacKinnon for his work on the structure and mechanism of potassium channels. Aquaporins are "the plumbing system for cells," said Agre. Every cell is primarily water. "But the water doesn’t just sit in the cell, it moves through it in a very organized way. The process occurs rapidly in tissues that have these aquaporins or water channels." There are four Aquaporins; Aquaporin 1, 2, 3 and 4. Aquaporins in plants classified into five subfamilies; Plasma membrane Intrinsic Protein (PIP) Tonoplast Intrinsic Protein (TIP), Nodulin-26 like Intrinsic Protein (NIP)-Small basic Intrinsic Protein (SIP), X Intrinsic Protein (XIP).
Aquaporins selectively conduct water molecules in and out of the cell, while preventing the passage of ions and other solutes. Also known as water channels, aquaporins are integral membrane pore proteins. Some of them, known as aquaglyceroporins, also transport other small uncharged solutes, such as glycerol, CO2, ammonia and urea across the membrane, depending on the size of the pore. For example, the aquaporin 3 channel has a pore width of 8-10 Ångströms and allows the passage of hydrophilic molecules ranging between 150-200 Da. However, the water pores are completely impermeable to charged species, such as protons, a property critical for the conservation of the membrane's electrochemical potential difference.
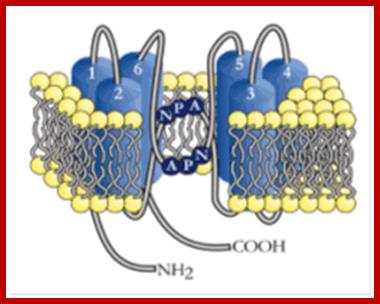
Plant Physiology and Molecular Biology; Water transport in plants; http://www.disafa-international.unito.it/
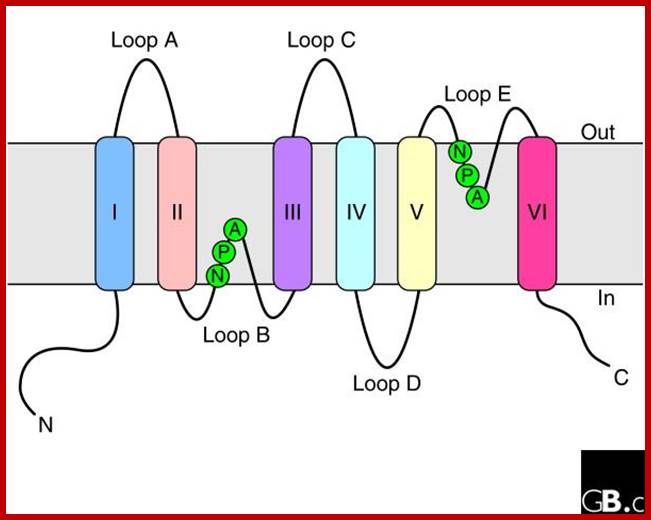
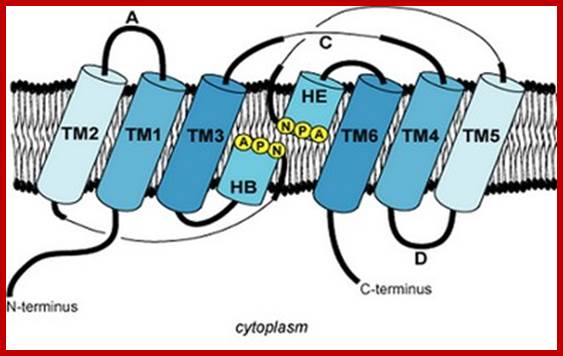
Topology of an aquaporin protein within the membrane. The protein consists of six transmembrane helices (I-VI) connected by five loops (A-E) and includes two internal tandem repeats (I-III and IV-VI, respectively). Loops B and E, containing the conserved NPA motifs (in the single-letter amino-acid code), form short α helices that fold back into the membrane from opposite sides. C, carboxyl terminus; N, amino terminus. http://www.genomebiology.com/
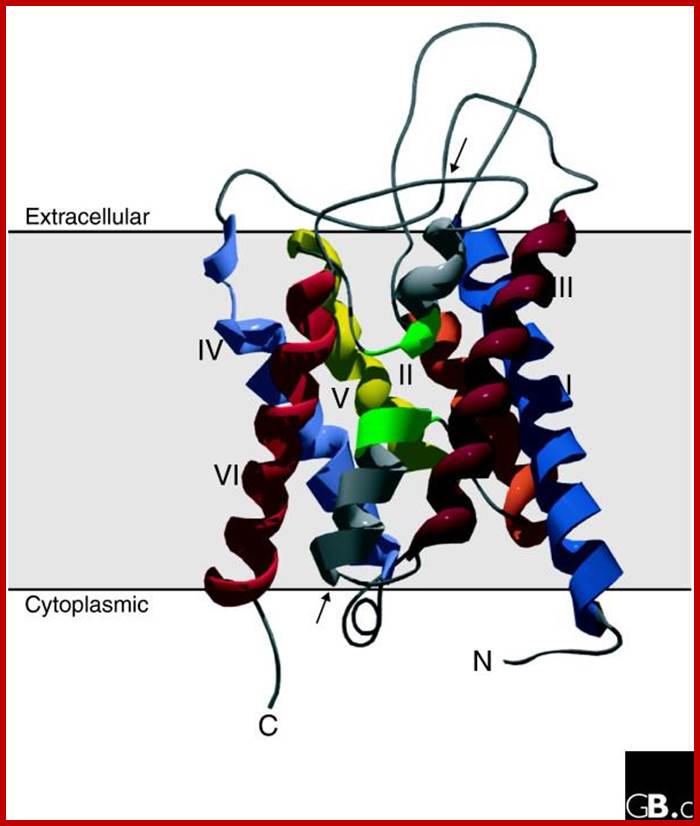
Three-dimensional structure of an aquaporin subunit monomer (a ribbon model of NtAQP1, a PIP1 protein from tobacco). The structure shows six tilted membrane-spanning helices (I-VI) and two pore-forming domains made up of two short α helices entering the membrane from the extracellular and intracellular surfaces (arrows). The two NPA boxes are indicated in green. Amino- and carboxy-terminal domains are oriented to the cytoplasmic side of the membrane. The figure was generated using MODELLER7v7 and Swiss-Pdb Viewer; This 'hourglass model' has been confirmed by three-dimensional maps of AQP1 using cryoelectronic microscopy. These maps also showed that aquaporins have a tetrameric organization: the four subunits are arranged in parallel, forming a fifth pore in the center of the tetramer. It is generally accepted that all aquaporin-like proteins assemble into tetramers. Each monomer alone can facilitate water flow, however. Recent experiments have indicated conductance of ions (K+, Cs+, Na+ and tetramethylammonium) through the central pore of the AQP1 tetramer; http://www.genomebiology.com/
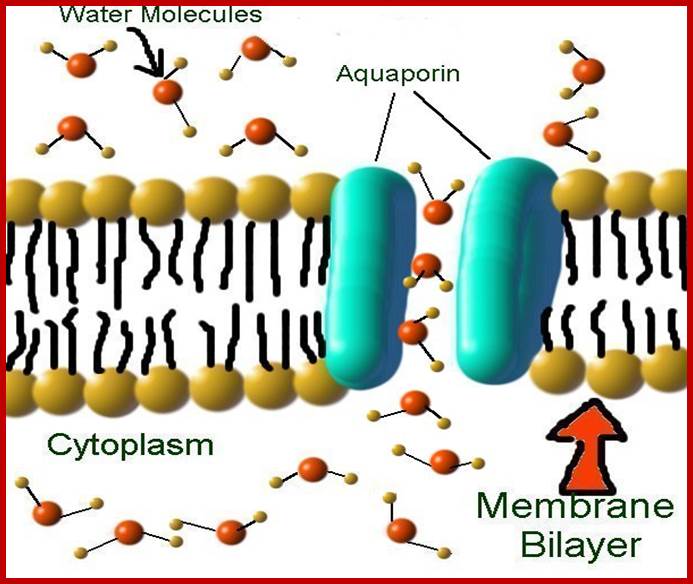 Aquaporins are water-specific pores that account for most of the
passage of water across biological membranes. Aquaporins can be
controlled indirectly as a blue light response. Aquaporins can be closed
by a phosphorylation switch. That is, the plant can control aquaporins by
using enzymes in a signal cascade. In this way, the plant can perceive a
stressful situation and hold on to its water; LSwatzell@semo.edu;http://cstl-csm.semo.edu/
Aquaporins are water-specific pores that account for most of the
passage of water across biological membranes. Aquaporins can be
controlled indirectly as a blue light response. Aquaporins can be closed
by a phosphorylation switch. That is, the plant can control aquaporins by
using enzymes in a signal cascade. In this way, the plant can perceive a
stressful situation and hold on to its water; LSwatzell@semo.edu;http://cstl-csm.semo.edu/
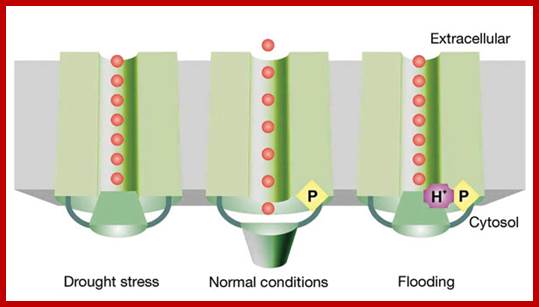
Structural mechanism of plant AQPs gating. Aquaporins act differentially under drought, flooding and under normal conditions. During drought stress the PIPs close in response to the dephosphorylation of two highly conserved serine residues (Ser 115 and Ser 274 of SoPIP2;1), whereas during flooding they close in response to the protonation of a fully conserved histidine (His 193 of SoPIP2;1). Susanna Törnroth-Horsefield et al; http://www.nature.com/
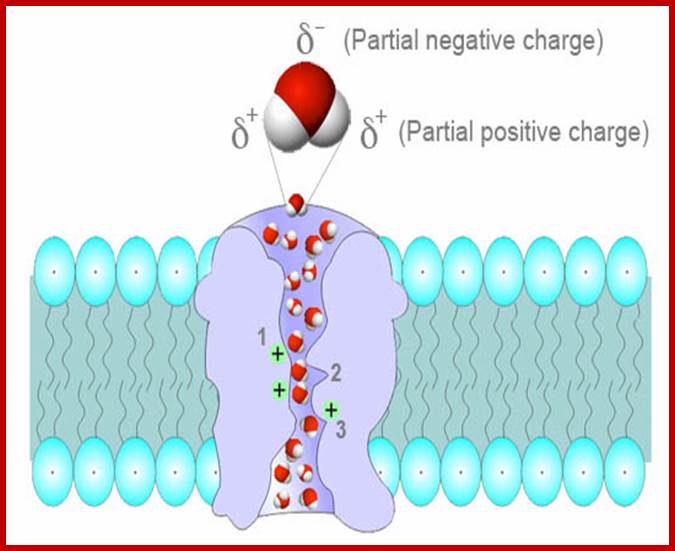
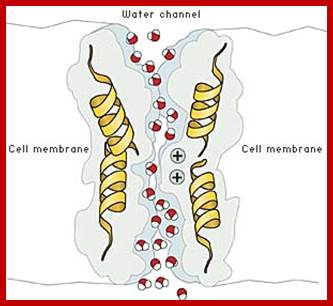
Cells have water channels; Aquaporin AQP structural features; helix-loop helix; TM domains produce transporter channel or pore for water transport. Water molecules are polar; the channel at the inner face has three positive charged sites, which helps in binding of negative ends of water molecules and transported across; askabiologist.asu.edu; www.biomedic.cl; http://en.wikipedia.org/asknature.com; http://www.uclouvain.be/
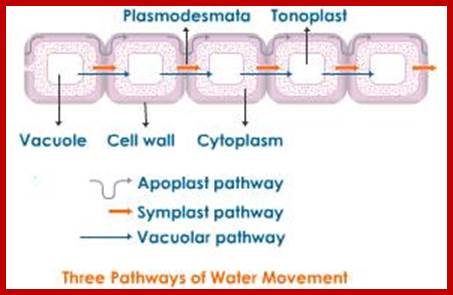
Once the water diffuses into cell walls and fills up the inter cellular spaces then water also diffuses into protoplasm, and thus water can move in the protoplasm from cell to cell through protoplasmic strands, plasma membrane and bulk flow across aquaporins provided the cells are living. There are possibly 36 aquapoin genes in plants identified by their (ESTs) cDNAs, but there are subtle differences between monocots and dicots Water is transported across aquaporins. The rate of movement through the protoplasm is extremely slow. Nevertheless, protoplasm to protoplasm movement is referred to as symplastic movement. Rarely one, finds the movement of water through the cellular vacuoles. The most rapid form of movement is Apoplastic and partially Symplastic.
Different models of AQPs are found in different organs in animal system; for example, Kidneys contain AQP1, salivary glands contain AQP3 and Interstitials contain AQP5. In plants there AQPs in both plasma membranes PIPs and Tonoplast membranes TIPs. The SIPs are found located in ER membranes. Overexpression of AQPs of Arabidopsis in tobacco plants resulted in increased growth, is deemed to be the effect of water uptake and photosynthesis. Aquaporin’s have a role in leaf movement. AQPs not only transport water but also soluble solutes across the membranes. Some AQPs such as AQP1 facilitate movement of CO2 and NH3.
Given that all aquaporins are structurally related and have highly similar consensus regions, particularly in the pore-forming domains, a similar transport mechanism can be assumed. The hydrophobic domain created by the loops B and E (Figure 2) has been suggested to be involved in substrate specificity and/or size restriction. The pathway through the aquaporin monomer is lined with conserved hydrophobic residues that permit rapid transport of water in the form of a single-file hydrogen-bonded chain of water molecules. The pore contains two constriction sites: an aromatic region comprising a conserved arginine residue (Arg195) forms the narrowest part of the pore, and the highly conserved NPA motifs form a second filter, where single water molecules interact with the two asparagine side chains]. Because of a direct interaction between water molecules and the NPA motifs, the dipolar water molecule rotates 180 degrees during passage through the pore. Both filter regions build up electrostatic barriers, which prevent the permeation of protons. In human AQP1, a hydrophobic phenylalanine side chain (Phe24) intrudes into the pore and enhances the interaction of single permeating water molecules with the NPA loops. In the bacterial glycerol facilitator GlpF, this residue is replaced by the smaller amino acid leucine (Leu21). Phe24 acts as a size-exclusion filter, preventing the passage of larger molecules such as glycerol through AQP1; Genome Biol.com
If conditions are favorable for absorption of water, one can visualize the pathway of movement of water from the soil solution into the vascular elements. First water enters into the AFS of root hair cell wall and then enters into the protoplasm. Bulk of the water absorbed by the root system enters through billions of root hairs which are continuously produced by the growing roots. Once water finds its way into such epidermal cells, it moves rapidly along the AFS spaces found in the cortical cells towards central vascular cylinder. But endodermal cells with their radial casparian thickenings are known to resist the free movement of water along the gradient. However, some of the endodermal cells located opposite to protoxylem elements are found to be free from casparian thickening and they act as free passage cells. Thus water ultimately finds its way into xylem elements.
MECHANISM OF ABSORPTION OF WATER
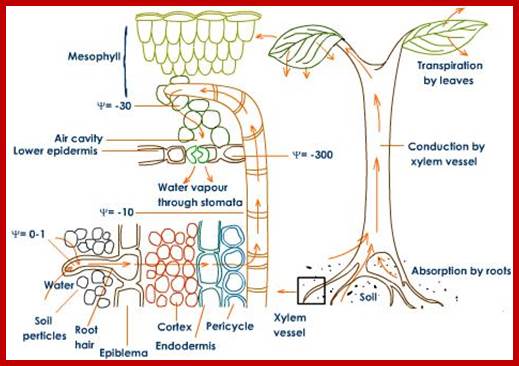
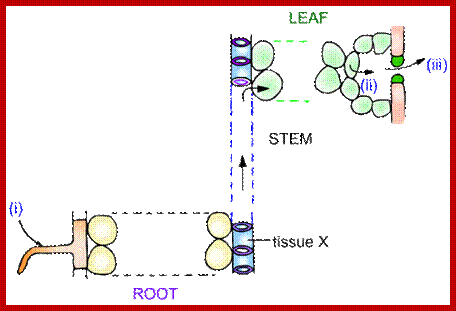
Various theories have been proposed to explain the mechanism of the absorption of water. Though a large number of plant physiologists are in the opinion that the process of absorption of water is by a passive mechanism, there are some, who still believe that active process also operates along with passive process, where passive process dominates over the active process. Passive is just water flow- from high concentration to lower concentration, further flow is an active process.
PASSIVE MECHANISM
Any physiological process that does not utilize metabolic energy is called passive process. The basic criteria for explaining the process of absorption of water in a passive mechanism, stems from the fact that there always exists a steep water potential gradient between the soil water and the cells including xylem elements found in the root system.
http://www.biology-pages.info
This gradient is always maintained because rapidly transpiring surfaces in the aerial regions develop a powerful transpiration pull which physically sucks the water upwards from the central vascular xylem elements. Thus, xylem elements in the root always experience a negative pressure, which is transmitted across the cortical cells towards the peripheral root hairs, which actually act as the absorptive surfaces. As a consequence of negative pressure operating in the central vascular xylem elements, water from the soil just diffuses rapidly along the gradient into the roots. As the transpiration pull is continuously operating the negative pressure is constantly maintained at the xylem elements. This greatly felicitates the rapid absorption of water by the root system. So the forces that are responsible for absorption of water generate not within the root system per se but in the aerial transpiring structures. More than 95% of the water absorbed by the root system is operated by passive forces like DPD gradient created by the transpiration pull. Aquaporins play an important role bulk flow of water across root hair to root cells.
ACTIVE PROCESS
Any process that requires the input of metabolic energy is called active process. In the absence of ATP or any other energy rich compounds such processes does not operate. One of the main reasons as to why some physiologists believe that active process is also involved in the absorption of water is because under favorable conditions, the roots of certain plants develop hydrostatic pressure within them. Such pressure is called root pressure. For example, plants like potato, tomato ad colochasia develop root pressure when there is plant of water is the soil probably due to heavy rains and minimum transpiration, due to high RH in the atmosphere. Under such conditions roots absorb more water than it can hold. As a result of excess of intake of water, a hydrostatic pressure builds up within the root. The development of root pressure in plants has been demonstrated in various species; where the bleeding of sap and even guttation has been attributed to root pressure. If respiratory poisons are provided root pressure does not develop. This is another evidence to show and that active absorption is responsible for the development of root pressure. It is very important to remember that active absorption need not lead to root pressure always. However, the mechanism of active absorption has been explained by two possible mechanisms.
1. Osmotically Active Mechanism: Under favorable conditions, as mentioned above, the special meristematic cells of the roots absorb and accumulate mineral nutrients from the soil. The same is actively transported across the cells towards xylem elements, into which nutrients are finally loaded. Absorption of minerals, transport and loading xylem is active processes, which require sufficient amount of ATP energy. As the xylem cells are loaded with more and more of minerals, a DPD gradient is created between the soil solution and xylem sap. This acts as the motive force for the movement of water into roots by passive osmosis. In this process, uptake of water does not require energy, but the accumulation of mineral salts which generates the force for the uptake of water, requires metabolic energy. Hence, this process is called osmotically active process. It is also believed that this process is mainly responsible for the development of root pressure. Interestingly, there is a good correlation between the conditions at which the root pressure develops and conditions at which nutrient uptake increases. It is very well known that roots absorb more ions and with a greater rapidity when a nutrient solution is very dilute. As respiratory poisons like DNP, inhibit ATP synthesis, they also inhibit active uptake of ions, thus inhibit osmotically active absorption and also the development of root pressure mechanism.
NON-OSMOTICALLY ACTIVE MECHANISM
Protagonists of this theory believe that the root pressure develops due to the uptake of water against its own concentration gradient. It is speculated that water is actually pumped into root cells by certain energy dependent pumps located in plasma membranes. Experimentally this hypothesis cannot be tested, because use of respiratory poisons also affects most of the energy dependent metabolic process including the active uptake of mineral ions, which is mainly responsible for the uptake of water by osmotically active process. Hence this theory has remained as an untestable theory.