PLANT GENETICS:
Plant_Cell_Genetics2-Genetics
Mendelian Genetics:

Gregor Johann Mendel
Mendel's life and work: Gregor John Mendel was born in 1822, to a poor farmer in Heinzendorf, now located in Czechoslovakia. The young Mendel was put to education; though he was-an ardent and sincere student he could not continue his education, because of poverty, so he gave up education and became a priest. As a monk, he had lot of work to do; occasionally he was called to do the job of stop-gap teachers. His interest in science and teaching had led him to Vienna, where he studied Physics, Mathematics and Biology in the local university. His academic career, though not brilliant, his training was of a great value.
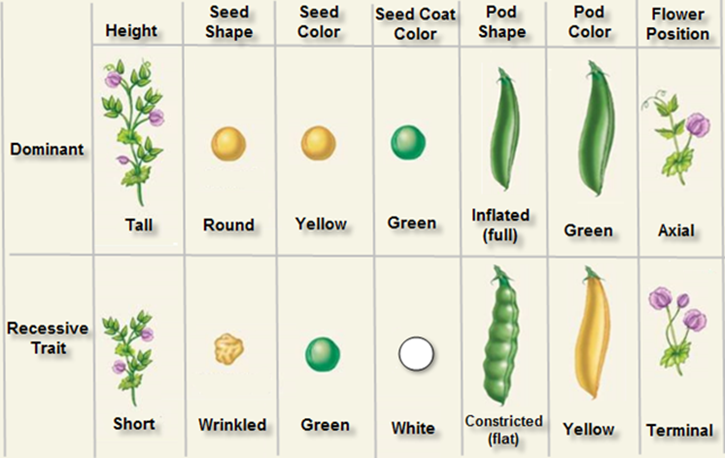
Upon his return to monastery in Brunn, he took up teaching profession seriously. But the young Mendel unsatiated with just teaching, was planning for experimental research. He had neither sophisticated tools nor the laboratory. The backyard of the monastery and garden pea plants has become his laboratory and tools. Paradoxically he selected garden Pea, what made him to select Pisum sativum plants as his experimental materials and collected 36 different varieties of Pisum sativum, which were different in their characters, of which he finally chose seven characters; what made him to select these seven characters is a mystery; which were distinct and showed clear cut variation. ex: Height- Tall & dwarf, color of the flower- violet & white. Position of the flower- axillary or terminal, color of the pod-yellow and green, shape of the pod-inflated and constricted, shape of the seed-round and wrinkled, color of the seed-yellow and green. To begin with he grew them separately and allowed them to get self-pollinated in successive generation. Finally he selected pure lines of these varieties (Pure line means a plant which on self-breeding produces the same kind). Then be grew these varieties of pea plants in separate plots, to prevent cross pollination. When plants grew and started flowering, he cross pollinated the flowers of desired characters for example, he cross pollinated round seeded varieties with wrinkled varieties; any of the parent plants can be taken as the acceptor. Using tweezers young flowers were opened and the stamens were removed. This process is called emasculation. Then pollen grains were collected from the donor variety and dusted on to the stigmatic surface of the emasculated flowers, and covered the artificially pollinated flowers with paper bags in order to prevent any foreign pollen contamination. Thus he crossed Tall with Dwarf, Violet with white, Terminal with Axillary, and Round with Wrinkled and so on. These experiments have to be done with care and skill. It is a hard job, but he did it meticulously and spent his time in the field from dawn to dusk. More than anything else he wrote everything he- did or observed in his diary.
Mendel in 1853 and 1963 conducted experiments on garden peas–Pisum sativum in the backyard of his church, there he cultivated ~5000 pea plants there he selected seven pairs of contrasting characters, then he crossed these varieties considering one pair of contrasting character, F1 allowed to self-pollinate and F2 also allowed to self-pollinate. What made him to select Pea plants and selected many phenotypic characters but concentrated on seven of the said characters for his experiments. This he did it without any knowledge of chromosomes or genes. His papers were published in Versuche über Pflanzen-Hybriden (Experiments on Plant Hybridization), in 1866. In 1900 his work was rediscovered by Hugo de Vries, Carl Correns, and Erich von Tschermak. Though Mendel’s work, at that time was little controversial, however the term ‘Genetics’ and ‘Alleles” were coined by William Bateson

Mother of all Genetics- “The DNA”
Mendel has chosen the above phenotypes and hybridize them and got the results called Mendelian Laws; www.blog.scs.sk.ca

Purple vs white;www.newtonsapple.org.uk
http://bioapbykelsey.wikispaces.com/
After completing cross pollination work in shortest possible time, he tendered the plants with care and love. The fruits were set, and started to fill. The excitement had grown in him. Probably he would have dreamt of all sorts of variations and combinations of his results of labor. However when the time came for harvesting, he plucked the ripened fruits, put them in separate bags and tagged them with names and analyzed the results.
Finally, he published his findings in a Biological Journal in which he explained - 1) the unit characters, as the factors of inheritance, 2) existence of them in alternate forms called alleles, 3) their transmission through gametes as pure forms, 4) their segregation in equal ratios, 5) their dominance' over recessive traits 6) independent but random assortment of factors, 7) their inheritance in a regular pattern in successive generation in set laws. But later he drifted towards other experimental materials like Hieracleum (a compositae member) and others. Unfortunately the materials were unsuitable and the results were quite contrary to the previous findings. He could not get anywhere. He started to doubt about his own findings and he was a distraught man. He was overburdened; he over worked; in this kind of stress and strains he passed away as any other mortal.
His work, though read and published (Ann. Proceedings of the Natural History society of Brunn, 1866), remained unnoticed or unrecognized for more than 36 years. However, in 1900, three biologists, Hugo Devries, Tschermak and Correns, while publishing their independent works on inheritance found Mendel's paper and to their surprise they realized that Mendel had already discovered of what they wanted or considered to be an important discovery. Thus Mendel's work was rediscovered, since then Mendel's name was etched, in the history of experimental biological science, in Golden letters, as the “Father of Genetics". Mendel was lucky in selecting the lucky seven characters in pea plants for one thing and for the other he was a priest, a churchman.
Snap peas= Pisum sativum:
TT and Tt- LE / le 2, gene, Gibberellin 3 beta-hydroxylase (converts GA20 to GA1), gibberellin 3-oxidases- PsGA2ox2; converts GA20 to active GA1; LE- stem length, GA biosynthesis GA1/ le dwarf. LE is the second gene to be cloned. The le is due to single base substitution or deletion
Flower color— RR and Rr- First gene to be cloned; it was a transcription factor
A1 and A2- A determine Anthocyanin pigment;
The A gene encodes a bHLH transcription factor. G to A transition in a splice donor site misplaced, so white color rr; “A’ is expressed throughout the plant;
Cotyledon color: ‘I’ for cotyledon color, I / i (SGR spod color stands forpecies including Rice), even senescing leaves remain green; this was due to reduced breakdown of chlorophyll.
Seed shape, RR = Rugosus and (Rr), R’ Is a regulatory gene, starch branching enzyme missing, SBE1.
Seed coat color; It has pleotropic effect. A and a2, - associated with flower color. Eurasian origin, mutation from G to A; splice donor site; require A and A2 required for the expression of Calchone synthase CHS (require 3 genes); bHLH gene is defective
Pod color, GP Green and gp yellow
Pod Shape; Pv, pv; suspected that P and V are linked; or V and LE linked; wild type inflated with sclerenchyma on the inner surface of the pod fruit wall. Mutants p and v responsible for constricted pods.
FA- Fasciation,
Position of flowers; Terminal or axial; FA wild type- linkage group IV which is syntenic. JI5= fasciation (Fa)
Most of the above mentioned Mendelian genes have been cloned or sequenced some and used for plant transformation experiments.
Mendel’s Laws of inheritance
Experiments with garden pea plants have made Mendel to lay down some principles of inheritance, which are now called as Mendel’s Laws of inheritance; the laws are. These laws were laid down without the knowledge of chromosomes, and gene linkage, segregation chromosomes and independent assortment and synteny of genes.
1. Law of unit characters
Every organism has certain distinct morphological as well as physiological characters. These characters are controlled by specific units of heredity. These were termed as factors. Now they are called as Genes. Genes exist in two alternate forms called alleles. Now it is known that the genes are located in chromosomes. For example, the color of the flower: Some plants produce red flowers and some white flowers. So the genes responsible for producing red color and white color must be different. As both the genes control the same character namely the color of the flower, they are called alleles or allelomorphs.
2. Law of Purity of gametes
The unit characters or hereditary units are transmitted to their offspring through gametes. During reproduction parents produce gametes which contain all the genes for all the characters of an organism. However, these genes are present in single doses. For example-pea plant may have hundreds of morphological and physiological characters. Among them, if we choose one character, then it is controlled by a particular pair of allelic genes. During reproduction only one gene enters into one gamete, thus gametes contain only one gene for this character. Thus the gamete is considered as pure and uncontaminated with respect to the said character. The same principle holds good for other characters also.
3. Law of dominance and Law of recessiveness
During reproduction, when a gamete carrying a particular gene fuses with another gamete carrying another gene belonging to the same pair of alleles, only one character is expressed in the offspring. For example, in pea plant when the gamete belonging to tall plant fuses with the gamete carrying 'dwarf' gene, the offspring 'sill have both the genes i.e. Tall as well as Dwarf, but only one is expressed; in this case it is the Tall character. Though the plant has 'dwarf' gene, only the Tall character is expressed because of the dominance of this character. Such genes are called dominant genes and the other unexpressed genes are called recessive genes.
In these cases, the morphological character that is expressed, that is visible, is called the Phenotype and the gene that is responsible for the expression of the phenotype is called the Genotype. For example, Tall character is called the Phenotype and the gene that is represented by a symbol, i.e., T, represents the Genotype. In Genetics, it is a convention to use the capital letter as the symbol of dominant gene; and small letter for the recessive gene. So T is dominant and this recessive and T and t form a pair of alleles. Furthermore, depending on the type of genes present, they are termed as dominant heterozygous (Tt), dominant homozygous (TT). But the recessive phenotype is expressed only when both the genes are homozygous recessive (i.e. tt)
4. Law of Segregation
A pair of allelic genes that is present in an organism for a given character, segregate in equal ratios during meiosis i.e. gametogenesis. This segregation of a pair of genes is just a chance and there is no choice for homologous chromosomes carrying these genes separate randomly. For example when the tall plant which is heterozygous produces gametes, only one (T) of the gene pair goes into one gamete and (t) to the other. In this process whether T gene goes to this gamete or that gamete is purely a chance process. However if thousand gametes are produced, 500 of them receive dominant "T" genes and the rest (500) receive "t" genes. Thus the segregation is random, but in equal ratios.
5. Law of Independent Assortment:
If two or more characters are taken together for observing the pattern of inheritance, different pairs of alleles behave independently and during segregation, they separate randomly as well as independently it because of random segregation of homologous pair of chromosomes. They are not linked to each other. This is because different pairs of alleles are located on different pairs of homologous chromosomes. While these genes express they do not influence each other and each of them remains independent and unique.
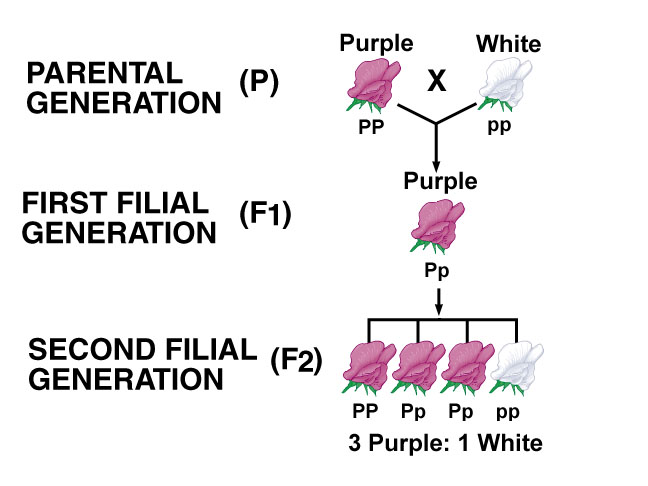
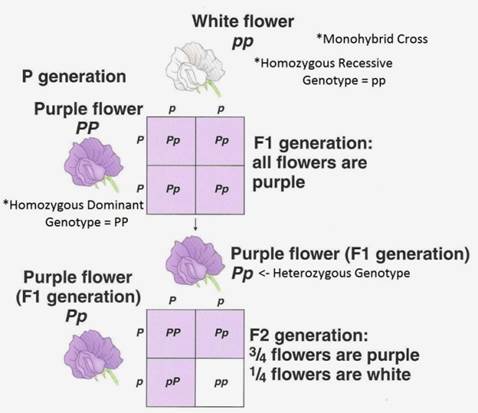
Monohybrid Cross:
1. One particular character which is controlled by a pair of alleles has been followed to find out the pattern of inheritance.
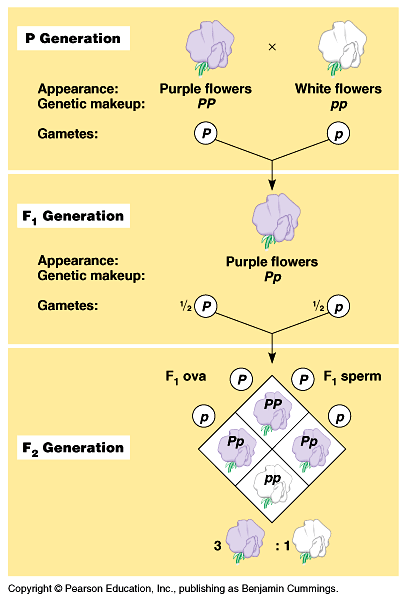
2. These pairs of alleles are located on a pair of homologous chromosomes. During gametogenesis these homologous pairs separate or segregate in equal ratios and each gamete receives one of the homologue pair, so also the genes. Gametes, with respect to a particular gene are pure, uncontaminated, and uninfluenced. In the FI generation only one character is expressed because of a particular gene, the gene that expresses the character is call dominant gene and the other which remains unexpressed is called recessive gene. Mating of the gametes is purely chance and random and not by choice. In the F2 generation the dominant recessive phenotypic characters are expressed in 3:1 ratio, but genotypically the segregation is 1: 2: 1. The statistical analysis of this type of inheritance is expressed as Monohybrid ratio.
Medelian Genetics; www.femitheist.net
Test Crossing or Back Crossing: In the case of Dominant Phenotype, by just looking at the morphology, it is difficult to find out" whether the given plant is heterozygous or homozygous for dominance. With regard to recessive, the character is expressed only when both the genes are recessive, thus the Phenotype also indicates the Genotype. In order to find the unknown Genotype of the dominant character, a cross breeding is performed which is called Test crossing or Back crossing. Here the dominant character is crossed with the known recessive character, for example Tall plant (its Genotype is not known) is crossed to dwarf (its Genotype is known). In the cross, if the off springs produced are of both dominant and recessive in equal ratio, the given plant is heterozygous for the said character. On the other hand if the offspring produced are all Tall, then the Tall plant is dominant homozygous. This type of crossing is employed to determine the heterozygosity or homozygosity of a suspected character.
Upon his return to monastery in Brunn, he took up teaching profession seriously. But the young Mendel unsatiated with just teaching were planning for experimental research. He had neither sophisticated tools nor the laboratory. The backyard of the monastery and garden pea plants has become his laboratory and tools. Paradoxically he selected garden Pea (Pisum sativum) plants as his experimental materials and collected 36 different varieties, which were different in their characters, of which he finally chose seven characters, which were distinct and showed clear cut variation. ex: Height- Tall & dwarf, color of the flower- violet & white. Position of the flower- axillary or terminal, color of the pod-yellow and green, shape of the pod-inflated & constricted, shape of the seed-round and wrinkled, color of the seed-yellow and green. To begin with he grew them separately and allowed them to get self-pollinated in successive generation. Finally he selected pure lines of these varieties (Pure line means a plant which on self-breeding produces the same kind). Then be grew these varieties of pea plants in separate plots, to prevent cross pollination. When plants grew and started flowering, he cross pollinated the flowers of desired characters for example, he cross pollinated round seeded varieties with wrinkled varieties; any of the parent plants can be taken as the acceptor. Using tweezers young flowers were opened and the stamens were removed. This process is called emasculation. Then pollen grains were collected from the donor variety and dusted on to the stigmatic surface of the emasculated flowers, and covered the artificially pollinated flowers with paper bags in order to prevent any foreign pollen contamination. Thus he crossed Tall with Dwarf, Violet with white, Terminal with Axillary, and Round with Wrinkled and so on. These experiments have to be done with care and skill. It is a hard job, but he did it meticulously and spent his time in the field from dawn to dusk. More than anything else he wrote everything he- did or observed in his diary.
After completing cross pollination work in. shortest possible time, be tendered the plants with care and love. The fruits were set, and started to fill. The excitement had grown in him. Probably he would have dreamt of all sorts of variations and combinations of his results of labor. However when the time came for harvesting, he plucked the ripened fruits, put them in separate bags and tagged them with names and analyzed the results.
Finally he published his findings in a Biological Journal in which he explained - 1) the unit characters, as the factors of inheritance, 2) existence of them in alternate forms called alleles, 3) their transmission through gametes as pure forms, 4) their segregation in equal ratios, 5) their dominance' over recessive traits 6) independent but random assortment of factors, 7) their inheritance in a regular pattern in successive generation in set laws. But later he drifted towards other experimental materials like Hieracleum (a compositae member) and others. Unfortunately the materials were unsuitable and the results were quite contrary to the previous findings. He could not get anywhere. He started to doubt about his own findings and he was a distraught man. He was overburdened; he over worked; in this kind of stress and strains he passed away as any other mortal.
His work, though read and published (Ann. Proceedings of the Natural History society of Brunn, 1866), remained unnoticed or unrecognized for more than 36 years. However, in 1900, three biologists, Hugo Devries, Tschermak and Correns, while publishing their independent works on inheritance found Mendel's paper and to their surprise they realized that Mendel had already discovered of what they wanted or considered to be an important discovery. Thus Mendel's work was rediscovered, since then Mendel's name was etched, in the history of experimental biological science, in Golden letters, as the “Father of Genetics". Mendel was lucky in select in g the lucky seven characters in pea plants for one thing and for the other he was a priest, a churchman.
Mendel’s Laws of inheritance
Experiments with garden pea plants have made Mendel to lay down some principles of inheritance, which are now called as Mendel’s Laws of inheritance; the laws are.-
1. Law of unit characters
Every organism has certain distinct morphological as well as physiological characters. These characters are controlled by specific units of heredity. These were termed as factors. Now they are called as Genes. Genes exist in two alternate forms called alleles. Now it is known that the genes are located in chromosomes. For example, the color of the flower: Some plants produce red flowers and some white flowers. So the genes responsible for producing red color and white color must be different. As both the genes control the same character namely the color of the flower, they are called alleles or allelomorphs.
2. Law of Purity of gametes
The unit characters or hereditary units are transmitted to their offspring through gametes. During reproduction parents produce gametes which contain all the genes for all the characters of an organism. However, these genes are present in single doses. For example-pea plant may have hundreds of morphological and physiological characters. Among them, if we choose one character, then it is controlled by a particular pair of allelic genes. During reproduction only one gene enters into one gamete, thus gametes contain only one gene for this character. Thus the gamete is considered as pure and uncontaminated with respect to the said character. The same principle holds good for other characters also.
3. Law of dominance and Law of recessiveness
During reproduction, when a gamete carrying a particular gene fuses with another gamete carrying another gene belonging to the same pair of alleles, only one character is expressed in the offspring. For example, in pea plant when the gamete belonging to tall plant fuses with the gamete carrying 'dwarf' gene, the offspring 'sill have both the genes i.e. Tall as well as Dwarf, but only one is expressed; in this case it is the Tall character. Though the plant has 'dwarf' gene, only the Tall character is expressed because of the dominance of this character. Such genes are called dominant genes and the other unexpressed genes are called recessive genes.
In these cases, the morphological character that is expressed, that is visible, is called the Phenotype and the gene that is responsible for the expression of the phenotype is called the Genotype. For example, Tall character is called the Phenotype and the gene that is represented by a symbol, i.e., T, represents the Genotype. In Genetics, it is a convention to use the capital letter as the symbol of dominant gene; and small letter for the recessive gene. So T is dominant and this recessive and T and t form a pair of alleles. Furthermore, depending on the type of genes present, they are termed as dominant heterozygous (Tt), dominant homozygous (TT). But the recessive phenotype is expressed only when both the genes are homozygous recessive (i.e. tt)
4. Law of Segregation
A pair of allelic genes that are present in an organism for a given character, separate or segregate in equal ratios during gametogenesis. This segregation is just a chance and there is no choice. For example when the tall plant which is heterozygous produces gametes, only one (T) of the gene pair goes into one gamete and (t) to the other. In this process whether T gene goes to this gamete or that gamete is purely a chance process. However if thousand gametes are produced, 500 of them receive dominant "T" genes and the rest (500) receive "t" genes. Thus the segregation is random, but in equal ratios.
5. Law of Independent Assortment:
If two or more characters are taken together for observing the pattern of inheritance, different pairs of alleles behave independently and during segregation, they separate randomly as well as independently. They are not linked to each other. This is because different pairs of alleles (genes) are located on different pairs of homologous chromosomes. While these genes express they do not influence each other and each of them remains independent and unique. There are exceptions to this.
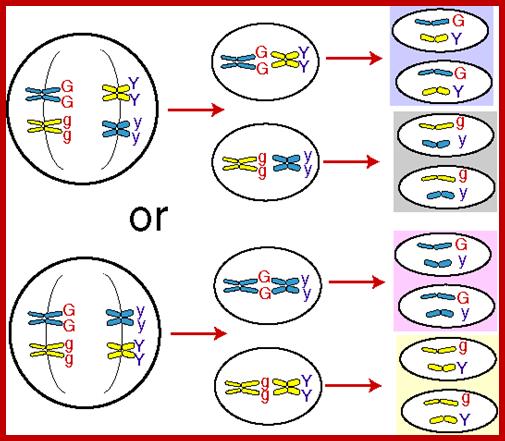
As homologous par of chromosomes separate equally but independent to each other we get the above ratio; Chromosomal basis of segregation and independent assortment;
hereditarygenetics.wordpress.com
Monohybrid Cross:
1. One particular character which is controlled by a pair of alleles has been followed to find out the pattern of inheritance.
2. These pairs of alleles are located on a pair of homologous chromosomes. During gametogenesis these homologous pairs separate or segregate in equal ratios and each gamete receives one of the homologue pair, so also the genes. Gametes, with respect to a particular gene are pure, uncontaminated, and uninfluenced. In the FI generation only one character is expressed because of a particular gene, the gene that expresses the character is call dominant gene and the other which remains unexpressed is called recessive gene. Mating of the gametes is purely chance and random and not by choice. In the F2 generation the dominant recessive phenotypic characters are expressed in 3:1 ratio, but genotypically the segregation is 1: 2: 1. The statistical analysis of this type of inheritance is expressed as Monohybrid ratio.
Test Crossing or Back Crossing: In the case of Dominant Phenotype, by just looking at the morphology, it is difficult to find out" whether the given plant is heterozygous or homozygous for dominance. With regard to recessive, the character is expressed only when both the genes are recessive, thus the Phenotype also indicates the Genotype. In order to find the unknown Genotype of the dominant character, a cross breeding is performed which is called Test crossing or Back crossing. Here the dominant character is crossed with the known recessive character, for example Tall plant (its Genotype is not known) is crossed to dwarf (its Genotype is known). In the cross, if the off springs produced are of both dominant and recessive in equal ratio, the given plant is heterozygous for the said character. On the other hand if the offspring produced are all Tall, then the Tall plant is dominant homozygous. This type of crossing is employed to determine the heterozygosity or homozygosity of a suspected character.
|
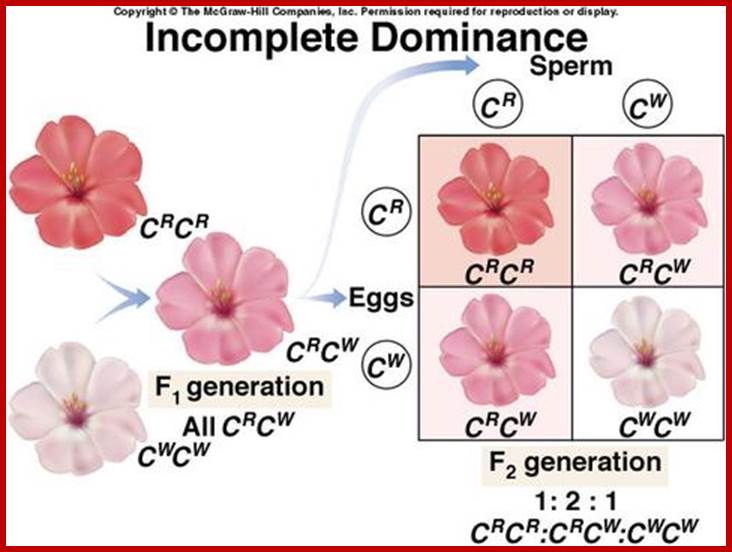
Incomplete dominance; show codominance, where both genes are equally expressed, showing both traits; a kind of blending expression. Several pairs of genes are involved here and thus produce intermediate color patterns; codominance would produce multicolored flowers. n; http://highered.mheducation.com/ |

Co-dominance in camellia cultivar; https://en.wikipedia.org/
In the case of colored Pisum flowers bearing red for both genes are functional, but in the case of heterozygocity, where one finds one dominant allele and another recessive; only dominant gene is expressed at molecular level and the recessive does not express anything and remain nonfunctional; such cases are called haplosufficiency. In another case where two genes are alleles one of the genes is normal, but another gene produces phenotype intermediate phenotype. In such cases the cause is due to haplo-insufficiency. In some functional allele in heterozygous state resembles the phenotype of non-functional-allele; this is another complete haplo-insufficiency.
Hardy-Weinberg principle (Estimation of carrier frequency)]:
Main article: Hardy-Weinberg principle
The frequency of
the homozygous state (which is the carrier state for a recessive trait) can be
estimated using the Hardy-Weinberg formula: ![]()
This formula applies to a gene with exactly two alleles and relates the frequencies of those alleles in a large population to the frequencies of their three genotypes in that population.
For example, if p is the frequency of allele A, and q is the frequency of allele a then the terms p2, 2pq, and q2 are the frequencies of the genotypes AA, Aa and aa respectively. Since the gene has only two alleles, all alleles must be either A or a and p + q = 1. Now, if Ais completely dominant to a then the frequency of the carrier genotype Aa cannot be directly observed (since it has the same traits as the homozygous genotype AA), however it can be estimated from the frequency of the recessive trait in the population, since this is the same as that of the homozygous genotype aa. i.e. the individual allele frequencies can be estimated: q = √f(aa), p = 1 − q, and from those the frequency of the carrier genotype can be derived: f(Aa) = 2pq.
This formula relies on a number of assumptions and an accurate estimate of the frequency of the recessive trait. In general, any real-world situation will deviate from these assumptions to some degree, introducing corresponding inaccuracies into the estimate. If the recessive trait is rare, then it will be hard to estimate its frequency accurately, as a very large sample size will be needed. Verbatum; https://en.wikipedia.org
Multiple Alleles:
A gene responsible for a particular function, in course of time may undergo a mutation and bring about a change in its function, hence in its phenotype. In a Population, it is possible that a wild type of gene may undergo mutation, resulting in multiple forms of the same gene. The phenomenon of gene existing in different forms is called multiple allelism or gene polymorphism. Such genes affect the phenotype in various ways. However, these genes may exhibit recessive, dominant or co dominant condition with the wild type gene.
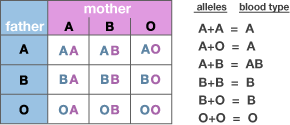
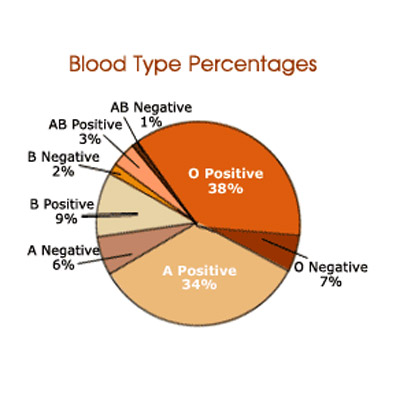
Blood Types and Their Genes; www.learn.genetics.utah.edu
Blood groups that children may inherit from their parents. https://en.wikipedia.org
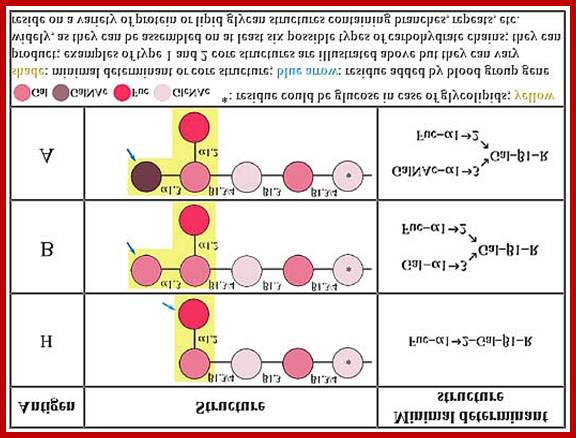
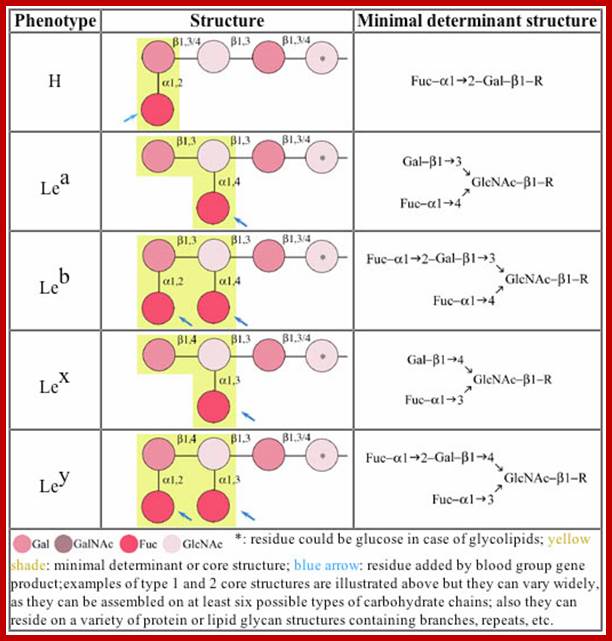
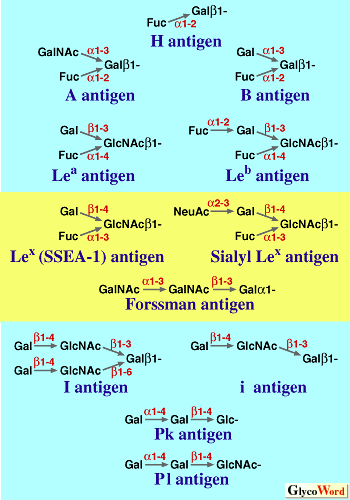
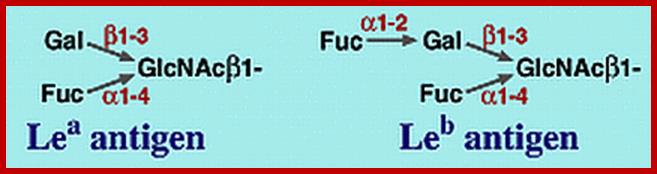
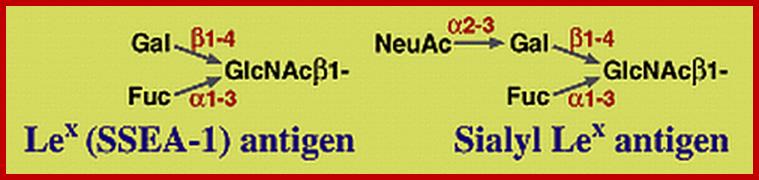
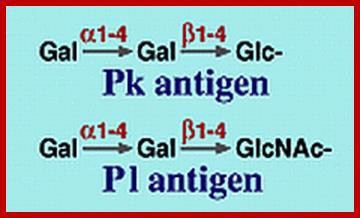
Lewis Blood Group Antigens; www.ncbi.nlm.nih.gov
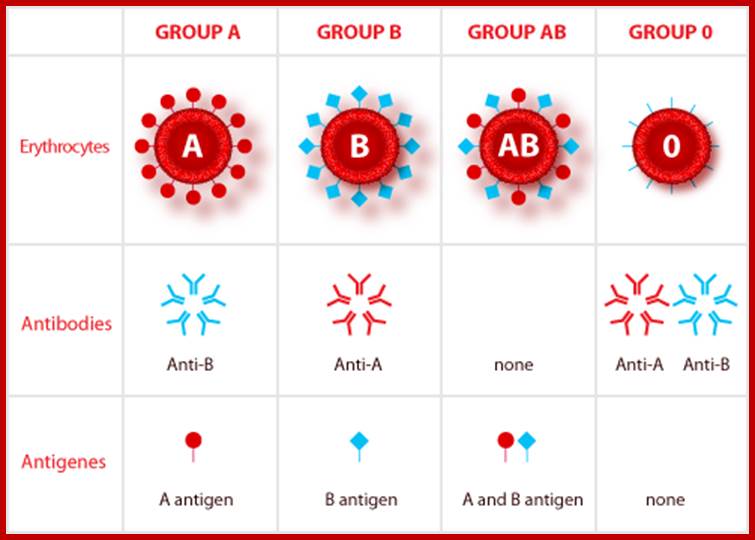
The existence of such multiple forms of genes in a population is not an uncommon feature, and the classical example is blood groups in human beings. Investigations in this regard have shown the presence of different types of blood groups, Rh+ Rh- and A.B.O. groups. Particularly A B O blood group is of immense importance for it is extensively used in blood transfusion in the case of accidents, severe anemia, or surgery etc. Landsteiner was the first to identify such blood groups. The variation in the blood groups is because of the membrane proteins of red blood cells (erythrocytes).
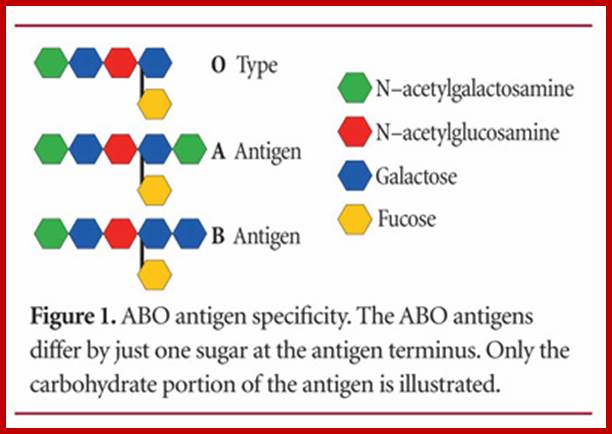
Some of the membrane proteins get glycosylated (addition of sugars) differently thus they produce various types of blood groups, thus they generate different antigenic property. Persons belonging to A blood group contain A type of glycosylated protein in the membrane of erythrocytes. (Similarly "B" and the "0" type). These biochemical phenotypes are due to the presence of A. B and 0 genes. The genotype of human beings may fall into any of the following types; AA, BB, 00, AB, AO or BO. In these cases the genes are codominant, it means both the genes express equally.
Identification of the blood types is very essential for blood transfusion, because, if by chance or mistake, blood of A type is given to B type person or vice-versa, the man who gets such blood type dies because of agglutination or aggregation of blood cells. This aggregation is due to antigen-antibody reaction.
Lewis and ABO blood group antigens:
A substance or substances, like proteins and polysaccharides etc., when injected into a human being, elicit or induce the synthesis of a class of serum proteins which react with the injected proteins and immobilize them. The substance that induces the synthesis of serum proteins is called an antigen (Agglutinogen) and the protein that is produced in response to antigen is called antibody (Agglutinin). It is a unique feature, that human beings are endowed with a capacity to produce specific antibodies against innumerable antigens. Based on this property of interaction between antigen and antibody, vaccines have been developed to combat various disease causing organisms like smallpox, tuberculosis, plague, cholera, polio; etc. Recently vaccine, against cancer also has been developed. Using antigen - antibody reactions blood types in human being can be identified.
Identification of Blood Groups:
It is always good to have blood group typed. Under emergency condition, it will be easier for doctors to perform blood transfusion without delay. In hospitals and other clinics, detecting the blood groups particularly, A, B and 0 types Rh + and Rh- and MN & S has been made very easy. The testing for the blood group can be performed anywhere if some basic facilities are available.
http://knowledgeisking.ning.com/
The best and surest way to detect is to have antiserum or antibodies against A, B, 0, Rh+ and Rh- antigens. Antibodies can be prepared by isolating antigens like A, B and Rh and the same if injected intra-dermally to an unimmunized Rabbit or Goat, the recipient reacts to the antigens and produces immunoglobulin proteins called antibodies against the said antigens. After second booster dose of antigen, antibodies can be collected by bleeding the animal. The blood obtained thus contains a variety of proteins; but the Immunoglobulins can be separated from the others by selective salt precipitation and column chromatographic procedures. Such purified antibodies against A, B, Rh+ antigens can be stored in deep freezers for a long time.
When testing for an unidentified blood type, few drops of blood is taken out of the person and same is applied on to four clean micro slides. Then a drop of dilute antibodies against A, B, O and Rh+ should be added to each of the blood drops on the micro slides. With the help of a glass rod they are mixed. Allow the antibodies and antigens to react for some time, and later the blood smears can be viewed under the microscope. Wherever cell aggregation or agglutination is observed, the blood type ca n be identified as A +, B' AB' Rh+. If agglutination is not found the blood of a person can be identified as A- (0), B-(O). O and Rh (-).
The principle of detection is simple. The antibodies raised against pure antigens, have certain specific regions in their protein called epitope recognition or binding sites, which recognize the antigens and they interact with them resulting in the binding between the two. When the blood of an untyped person is mixed with pure antibodies, the antigens present on the erythrocytes or blood cells interact with the antibodies and bring about agglutination of cells. If aggregation is detected in a particular serum, say A serum, the blood type can be identified as A. On the contrary if the blood cells do not agglutinate with A serum, but do so with B serum, then the blood type can be said to be as B. On the other hand, if the blood cells of that person show agglutination against both the A & B serum, it can be said that the blood group of that man is AB. Notwithstanding this, if there is no reaction against any of the A or B sera, it can be concluded that the blood is of 0 type.
With regard to Rh+, if the blood cells get agglutinated against Rh+ serum, the person is Rh+, if not he or she is Rh- negative R, Similarly any type of blood can be detected, if one possesses the right type of antibodies.
Wikimedia.org
ABO Blood; http://www.icr.org/
|
Blood cells of untyped persons |
Antibodies against |
Conclusions |
||
|
A |
B |
Rh+ |
||
|
1 |
+ |
- |
+ |
A type and Rh + |
|
2 |
- |
+ |
- |
B type and Rh - |
|
3 |
+ |
+ |
- |
AB type and Rh - |
|
4 |
- |
- |
+ |
O type and Rh + |
|
5 |
+ |
+ |
+ |
AB type and Rh + |
|
6 |
- |
- |
- |
O type and Rh - |
http://www.ncbi.nlm.nih.gov/
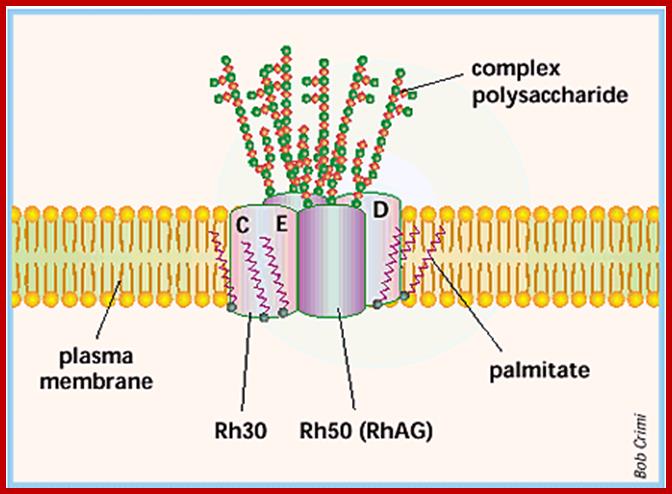
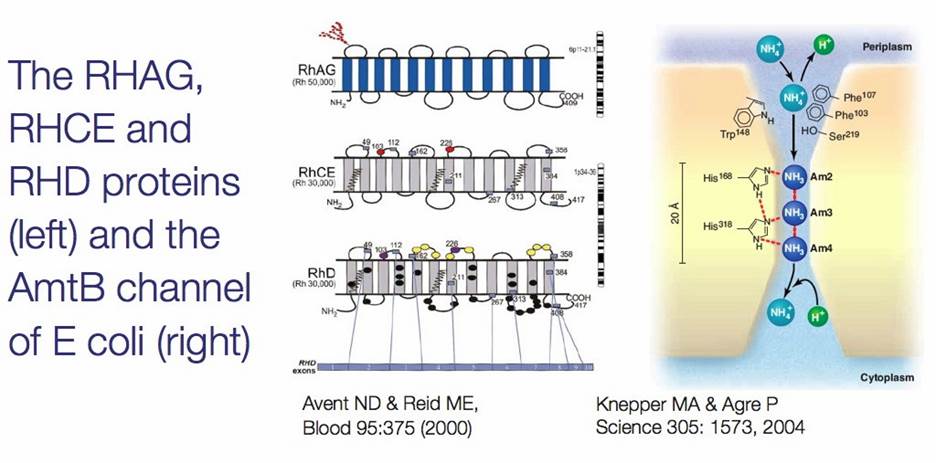
Rh group; www.ncbi.nlm.nih.gov
Blood groups.www.nfs.unipv.it
Similar to ABO type there are other characters which are controlled by multiple alleles. In humans MNS series have 20 multiple alleles, Kell series have 6 and Rh series have 30 or more alleles. Another significant aspect of blood group genotype is co dominance, where both the genes express their character equally without affecting the phenotypic character of the blood; Ex. A and B genes express their antigens equally and both remain in the same man without any interaction or modification.





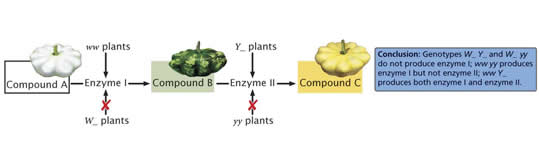
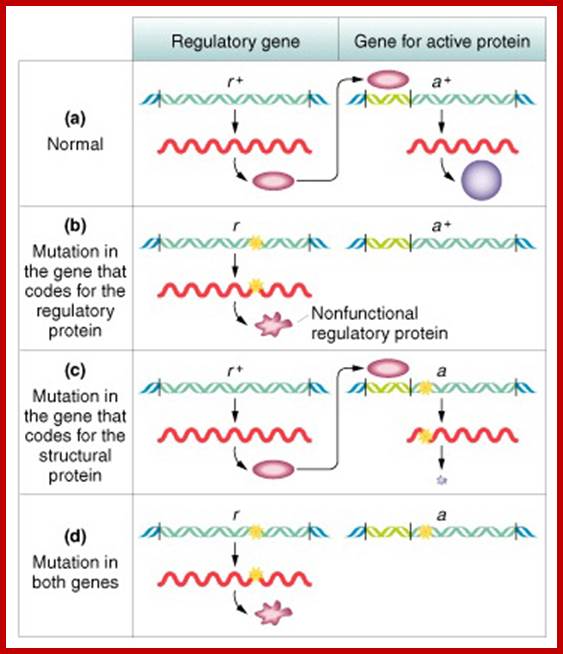
Epistasis: Masking mutants allele Effects) One gene, which is independent of the other, capable of suppressing the expression of the other gene, but it is not recessive and such a gene is called epistatic and the phenomenon is called epistasis where a gene product blocks the expression of the other gene, but both genes are independent of each other. If a mutant alleles exist for genes controlling two steps in the same biochemical pathway, recessive homozygotes for one gene may mask the effects of another. Interacting genes in same pathway with different mutant phenotypes may produce a\modified (9:3:3:1) ratio of 9:3:4. Epistasis points to interaction of genes in some biochemical or developmental sequence.
{kind=link}
{kind=link}
{kind=link}
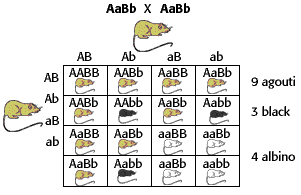
Priciples of genetics;
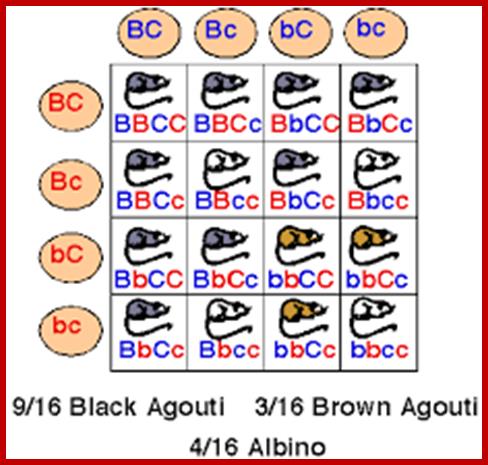
imgarcade.com;www.biposery.flu.edu

skin color in Labrador;; http://www.bio.miami.edu/
The B locus controls the color of melanin pigment: B is black; b is brown
The E locus prevents melanin from being fully deposited in the hair shaft: E is full pigment; e is blocked pigment
The E locus is epistatic to the B locus: It prevents the complete deposition of the already-made melanin pigment in the hair shaft (though it does not affect deposition in the skin).
Thus, the epistatic locus operates "developmentally downstream" (i.e., after the B locus is already expressed) from its hypostatic gene.
Possible genotypes/phenotypes:
- B- E- will be black (fur and skin)
- B- ee will be yellow fur, dark skin
- bb E- will be brown fur, brown skin
- bb ee will be yellow with brown skin
The golden coat is an example of recessive epistasis: the recessive allele of a gene (i.e., the e allele that prevents pigment deposition) exerts epistasis over another locus (the B locus, controlling pigment color), resulting in the pale-colored hair shafts. http://www.bio.miami.edu/

Pigments of carotinoid family are responsible for different colors on capsicum/chillies; Bell Peppers color:Gene interaction; http://www.bio.miami.edu/ https://www.quora.com
- Y - timing of chlorophyll elimination (Y - early; y - normal)
- R - color of carotenoids (R - red; r - yellow)
- C - regulation of carotenoid deposition (C - normal; c1, c2 - lowered concentration)
- This leads to a few possible genotypes producing interesting phenotypes:
- Y- rr c1c2 - pale yellow
- Y- rr Cc2 - darker yellow
- yy rr CC - green
- Y- R- CC - red
- yy Rr CC - purple
- Y- Rr Cc2 - pale yellow
www.imgarcade.com
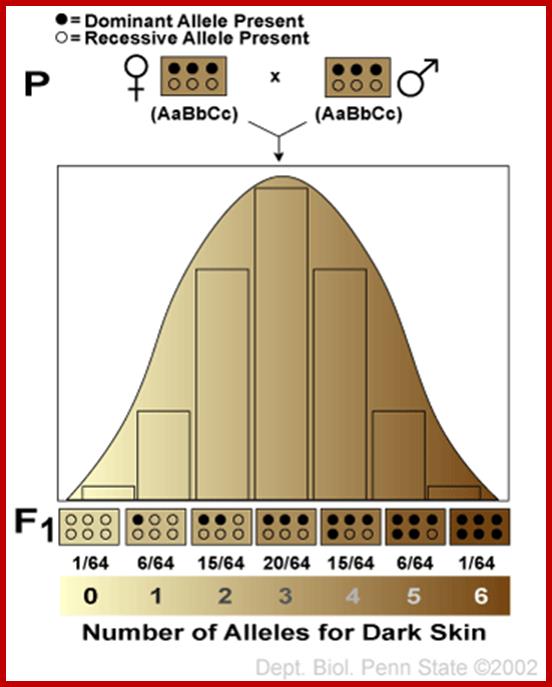
Multiple gene inheritance:
A particular phenotypic character is controlled by not a single gene but many affecting the same character. This type of inheritance is called multiple factor inheritance and quantitative. Examples- skin color and height in human beings.
www.imgarcade.com
Sex determination and sex linked inheritance:
Organisms with morphological sex difference as males and females, the sex of each of them is generally determined by sex chromosomes, which contain sex determining genes. Those chromosomes which determine sex of an organism are called sex chromosomes and other chromosomes are autosomes. In human beings there totally 2n = 46 chromosomes, of which 44 are called autosomes and the other two are called sex chromosomes, where they constitute two types, one is called X chromosome and the other is called Y chromosome. A male individual contain 2n=44A + XY and the Y is male determining chromosome, but the female contain 2n=44A + XX, where X is female determinant. This rule of XX and XY differs in different systems.
http://www.brusselsgenetics.be/
A majority of plants and animals exhibit sexual mode of reproduction. Some of them are bisexual (hermaphrodites) and some of them are unisexual. In higher animals unisex is a predominant feature, where male and female sexes arc distinguished by characteristic structural and functional features. Even many plants exhibit this kind of sexual dimorphism.
Sex chromosomes and Autosomes:
Cytogenetic studies, in the past fifty years or so, have established that the sex of on organism, in its early development is determined by special chromosomes called Sex chromosomes or Allosomes such as X and Y. All other chromosomes are called Autosomes (A). All unisexual animals including Insects, Fishes, Frogs, Birds and mammals including human beings show XY mechanism of sex determination. Even some unisexual plants like Melandrium, Coccinia, Sphearocarpus etc. show X/Y mechanism. However, certain variations are also found.
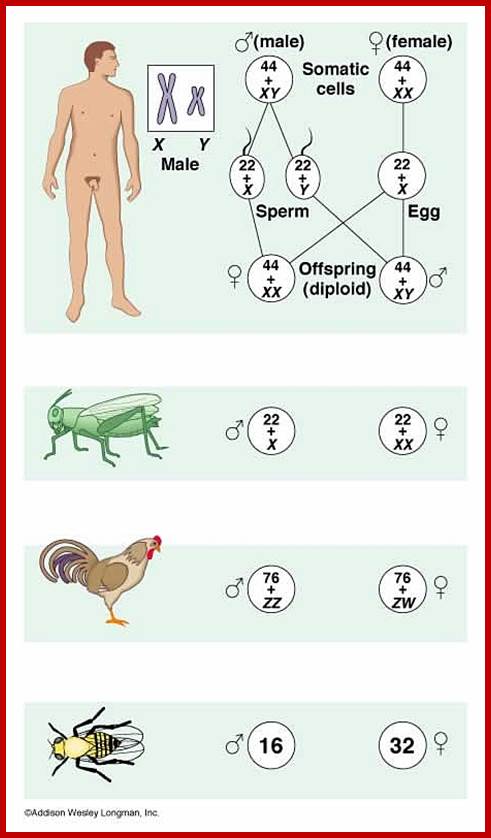
(1) Sex determination in human beings:
The diploid number of Homo sapiens is 46. In females, 2n = 44 A + XX is the chromosomal pattern, but in males 44A + XY is the chromosomal composition.
The X chromosome in males is euchromatic (genetically active) and Y chromosome .is completely heterochromatic; assumed to be genetically inactive. In fact, it is now established that the male determining gene is located in Y chromosomes and the female determining gene on X chromosomes.
There are cases where the chromosomal composition in certain patients was found to be 44A + XO i.e. one of the sex chromosome is missing and the sex is female. In another case 44A + XX shows male sex. This is because, in the first case me Y chromosome is completely missing. So the X chromosome determines the sex of an individual as female. But in the second case (44A + XX), a segment of Y chromosome containing male determining gene(s) is somehow translocated to one of the X chromosomes. In this special case, in spite of the absence of Y chromosome the sex of the patient was male, because the translocated segment of Y chromosome contained sex determining gene. This clearly establishes it is XX and XY combination that ultimately determines the female & male sexes respectively. The same is true with many higher animals and also some plants.
Out of two X chromosomes in females one is euchromatic and the other is total heterochromatic and it appears as a bar body in females. In males the Barr body is absent If by chance a Barr body is identified in males, then his chromosomal composition is 44A + XXY, and it is an abnormality.
But in birds 2A + ZZ determines male sex and 2A + ZW determines female sex.
2. Sex determination in Drosophila: In Drosophila, mosquitoes and other dipterans insects, the sex determination is by XX and XY mechanism, but with a difference when compared to the sex determination of human beings. Its 2n number is 8. If 2n = 6A +XX-it is female. If 2n = 6A+XY then it is male. But if an insect has 2n = 6A with only one X chromosome, then the insect develops as a male. This is due to the balance between autosomal number and sex chromosomes; here male sex is determined in spite of the absence of Y chromosomes.
6A+XX = Female 6A 4- XXY = Super female
6A+XY = male 6A + XYY = Super male
6A L XO = Male
Molecular mechanism of sex determination in Drosophila has been more or less elucidated. The genes involved in the processes are sXl, dsXl and few other related genes. The alternate splicing of the said gene transcripts plays a significant role in determining the sex of the organism at an early stage of tissue differentiation.
3. Sex determination in Honey bees: In honey bees, the queen bee lays eggs and the worker bees collect honey. The queen bee is always diploid and it alone is capable of laying eggs. If the eggs fertilize with a sperm, the animal develops into females. On other hand if the haploid egg remains unfertilized, it develops into a male donor bees or slave bees.
4. Sex determination in bacteria: In bacteria, males arc slightly bigger in their cell size than their female counterparts. The male sex of bacteria is determined by the presence of an episode containing fertility factor. If the said factor is absent, then the bacteria act as female.
SEX LINKED INHERITANCE
Similar to Autosomes, sex chromosomes particularly X & Y chromosomes also carry expressible genes.
As sex chromosomes determine the sex of an organism, the genes found on sex chromosomes, as a linkage unit, also express their phenotype which is restricted to a particular sex of an organism. Such type of inheritance and expression limited to only one or the other combinations of sex chromosomes is called sex linked inheritance. So certain characters are inherited to either female or male sexes.
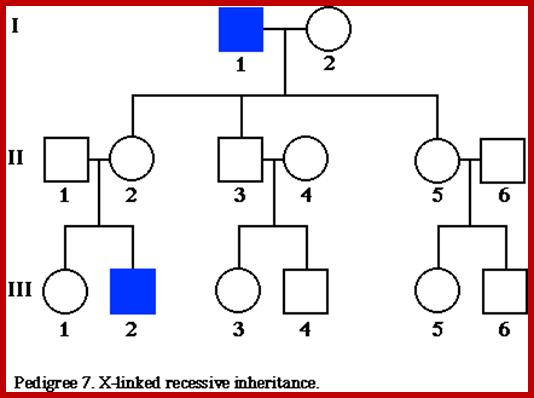
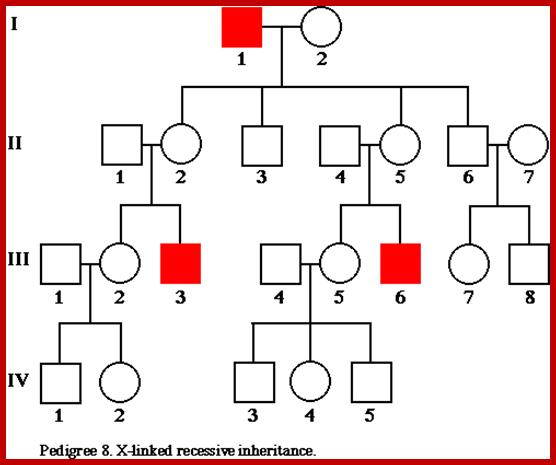
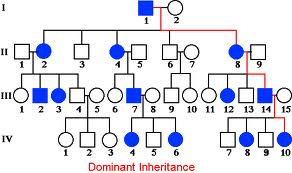
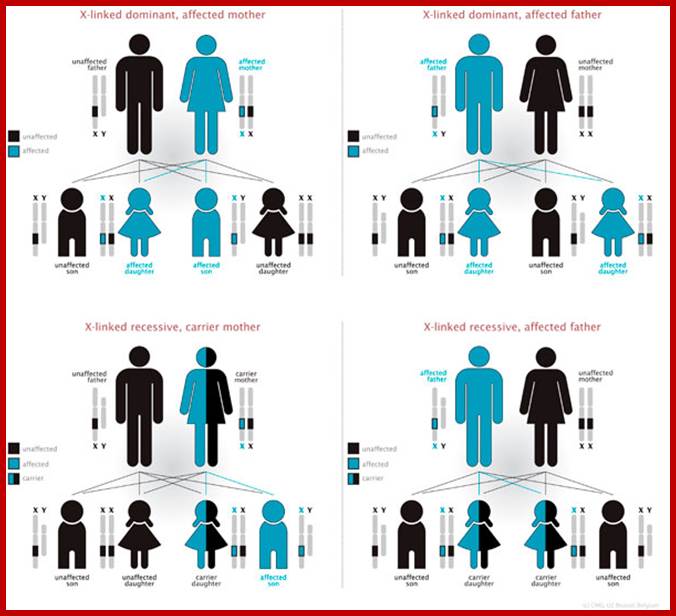
X-linked characters: Certain characters like color blindness, hemophilia muscular dystrophy in human beings, red and white eye color in Drosophila melanogaster; and black and barred plumage in poultry are some of the examples of sex linked, specifically X linked characters.
Y-linked characters: The Y chromosome also contains many genes in human beings but they don't express. Even if expressed, it is difficult to identify. But recent investigations do indicate that Y chromosomes also carry genes which express only under certain intracellular environment A gene for hypertrichosis i.e., hairs on external pinna, male sex determining factor, HLA - antigen and overall height controlling factors have been assigned to Y chromosomes.
PATTERN OF SEX LINKED INHERITANCE:
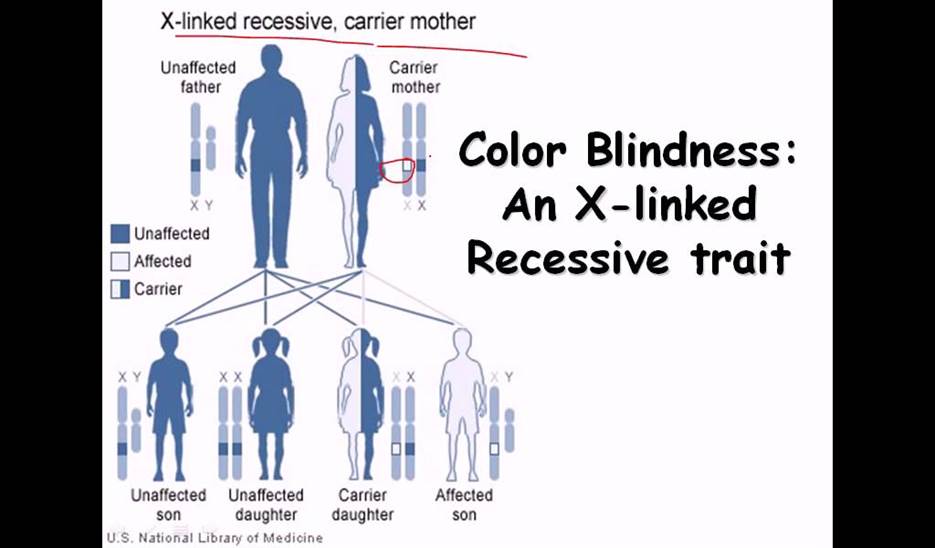
Eg. Color blindness:
Persons having color blind trait cannot distinguish red color from the green color. This character is controlled by a receive gene called “c”. The normal wild type is "C" gene. These genes arc located on X chromosomes. If the female is heterozygous for this character i.e., “Cc, her color sensitivity is normal. On the other hand, if she is homozygous for color blind genes “cc" she is color blind. On the other hand, if a male carries a "c” gene on X chromosome; he is color blind; it is because the Y chromosome docs not carry any normal allelic and it is genetically inactive.
Criss-cross inheritance: If the female, heterozygous (Cc) for color blind gene, marries a normal male, (CY), the probability of getting female to male children is 50:50. Among males, 50% will be color blind and other 50% normal. But all the females will be normal Follow the diagrammatic representation below.
Instead, if a heterozygous (carrier) female (Cc) marries a color blind male CY: 50% of the females and 50% of the male children will be color blind and other male and female children arc normal.
The same pattern of sex linked inheritance is also found with respect to hemophilia gene.
SEX INFLUENCED CHARACTERS:
Not all phenotypic characters expressed in females or males show sex-linked inheritance. For example, in human species beard, moustache, hair development on the chest, voice cracking, masculinity etc., are restricted to males. Similarly, non-development of beard, moustache, chest hairs, non-cracking of voice and development of breast etc are restricted to females. The above said characters, restricted to a particular sex, are expressed not at the early childhood but manifested at the adolescent stage. Such characters arc called sex influenced or secondary sexual characters.
The genes responsible for the expression of such characters are not found on sex chromosomes. However, the expression of such characters is under the influence of sex hormones and the age. Even masculinity and feminism are secondary sexual features.
Another good example is milk production in cattle. The genes responsible for milk production are present in both males and females, but they are expressed only in females because the female hormones influence the genes to produce milk.
Sex limited characters: Development of horns mostly in male sheep, formation of characteristic comb and feathers in male cocks and baldness mostly in male humane are actually sex limited characters.
GENETIC DISEASES
Heritable disorders are called Genetic diseases and they arc often lethal. Such disorders go unnoticed in plants, but in higher animals, especially in human beings, these diseases arc of great concern to the mankind. One should note that not all congenital diseases arc heritable; some of them arc acquired due to pathogens.
Genetic diseases are due to mutations. They may be induced or spontaneous. Mutation in single genes (point mutations) or changes in the chromosomal structure (aberrations) or in number (ploidy) leads to variations in the morphology and function. Some of these changes are inherited to off springs. The genetic diseases may express at birth itself or later.
In fact, Garrod and Landsteiner were responsible for the discovery of inborn errors in human beings. Since then, a large number of such diseases have been identified and the details of such diseases have been studied at the biochemical level and also at the molecular l~3Vel. The recent innovations in the field of genetic engineering technology have provided tools and techniques to identify such genetic disorders. In fact, in many western countries like Great Britain, West Germany, France and USA, facilities are available in many medical centers, for genetic screening and genetic counseling.
Some of the genetic diseases known to man are color blindness, Hemophilia, Phenylketonuria, Alkaptonuria, Albinism, Tyrosinosis, Brittle bone, Autoimmunity, Sickle cell anemia, Genetic dwarfism, Huntington's chorea, Erythroblastasis faetalis, Turner's syndrome, Down's syndrome, Klienfelter's syndrome, Thalassemia, Glactosemia, Diabetes, Alzheimer’s disease etc. Even families prone to heart disease and alcoholism have been traced to genetic disorders.
Phenylketonuria (pku): This disease is caused by point mutations. In this case the enzyme responsible for the conversion of phenyl pyruvate to hydroxypyruvate is inactive; hence phenyl pyruvate accumulates in the body. The result is the mental disorder and such persons are called Phenylketonuria idiots. However this disease can be kept under control by providing the substrates in the diet.
Alkaptonuria: This is another genetic disease caused by the disorder of enzymes responsible for converting Homogenistic acid to acetoacid. Because of this, patient's urine turns blue black as it comes in contact with air. This disorder also manifests in mental malfunctions. Again this disease can be controlled by feeding the patient with a diet containing the substrate.
Turner's syndrome: This is a chromosomal disorder is due to the loss of one X chromosome (2n = 44A + X). Patients exhibit deformities like enlarged genitals, short stature, and webbed neck, broad shield like chest, under developed breasts, uterus and ovaries. Their mental behavior is like idiots. They are female individuals and their life span is short. There is no cure.
Klienfelter’s Syndrome: The disease is caused by the presence of one extra X chromosome along with the normal XY chromosomes, (2n = 44A + XXY). The person afflicted with this kind of disease are males; long legged, develop female type of breasts, voice docs not crack, beard and chest hairs do not develop. In this case also there is no permanent cure.
Down's syndrome: Persons have one extra autosome (21st autosome). The chromosomal constitution is 2n = 44A + 21stA + XY. The person is male and exhibit mental disorders. Peculiar foldings of eyelids, short stature, stubby hands and feet, malformation of heart etc., these patients arc also called as Mongoloid idiots. There is no cure for this disease.
Huntington chorea: This disease expresses only at later stage of the life, say around 45-60 years of age. They suffer from extreme body tremors due to single gene defect this is also due to cavitations in the brain, which causes neural disorders.
Diabetes mellitus: In most of the cases, this disease is expressed at the age of 45-50 years (Diabetes II). The sugar level of blood rises very high due to the absence of functional insulin molecules or the insulin fails to stimulate sugar uptake by cells. High levels of blood sugar causes disorders in the body and in extreme cases patients go through comatic stage from which they may not recover at all. However, by proper intake of insulin one can lead a normal life. This affliction is global phenomenon, nearly 20- 30 percent of the population in developed countries have one or the other type of Diabetes; namely Diabetes-1 and Diabetes-2; where the former is juvenile and the latter is an adult disease.
Hemophilia: Hemophilia is often called bleeding to death disease. In normal humans a cut leading to bleeding stops with clotting of the blood. But in hemophiliacs, the defective gene does not produce clotting factor that is why they bleed to death. In hemophiliac there arc some variations, such as .external and internal bleeding. This disease is not autosomal linked, but sex linked.
Sickle cell anemia: This disease is very common in black -population of Africa. The normal hemoglobin gene produces functional oxygen carrying hemoglobin protein. In a mutant (just one amino acid substituted) the hemoglobin is rendered nonfunctional. In homozygous both genes are not functional. Hence the RBC cells collapse for the lack of sufficient oxygen, and the cells show sickle shape. People from this disease suffer from the lack of sufficient oxygen, and often gasp for the breath. But the heterozygous persons though show symptoms, they are immune against malarial infection.
Alzheimer*s disease : This disease manifests at old age and the diseased people suffer from forgetfulness ex, misplacing the key, leaving the key some place is not the disease, but having the key if one fails to unlock with the key is a clear manifestation of the disease. This has been attributed to single gene.
Persons suffering from such genetic diseases should go through proper diagnosis and treatment. This can be done through proper genetic counseling. Though there are no permanent cures, at least by proper care and precautions they can be maintained till they die a natural death.
Duchene Muscular Dystrophy: A membrane protein is involved in muscle contraction. Mutation in this gene can cause abnormality in muscle contraction. It is X-linked
HEREDITY AND VARIATION
Inheritance of parental characters by their off springs is referred to as Heredity. The material or chemical basis of heredity is DNA. Except for certain viruses where the hereditary material is RNA, all other organisms contain DNA as the hereditary material. However, all individual biochemical characters are controlled by unit factors of inheritance called Genes. In the classical concept of Mendelian genes all the genes were once considered as units of heredity, controlling specific characters; but; now they are considered differently. One gene: one character concept does not hold any waters today. A character, like red color of the flower, is not the product of one gene, but it is the ultimate expression of a series of biochemical steps which are controlled by different enzymes, If any one of the enzymes is missing or malfunctioning the ultimate character is not expressed. As the enzymes are the ultimate expressions of one or more genes, one gene: one character is no longer tenable.
Recent investigations into the molecular-nature of the gene have revealed more complex organization of the gene as a unit character. Though a specific segment of DNA is considered as the unit of function in terms of its expression, the fine structural analysis of the gene shows that the gene is not only split or spliced, but also overlapping.
A spilt gene is that which contains some segments of the DNA that are coding, called Exons, which are interspersed with non-coding regions called Introns. Notwithstanding this, the genes are overlapping, in the sense, that a number of base sequences which are sufficient enough for 6 proteins actually code for 9-10 proteins. This is possible only when the reading frame overlaps on another segment. This ultimately boils down to the vie w that the start and the end of the gene depends upon the recognition of the initiator sites which in turn are controlled by the cytoplasmic factors. However the concept of gene as unit of heredity, irrespective of spilt nature or overlapping of characters, is still valid in terms of Neo-Mendelian Genetics. Whatever may be the nature of a gene, it is the unit (consisting of a segment of DNA) which is capable of producing a meaningful polypeptide.
In molecular terms, a particular character is the result of a particular gene or a group of genes. If such gene or genes undergo changes at the DNA level, the ultimate character also gets changed. This change is referred to as variation. If such a change is heritable, it is called Mutation.
Mutation is possible at the chromosomal level or at the gene level. At the chromosomal level, the entire set of chromosomes may be involved and such a change with reference to the basic chromosomal set is called ploidy. It may be either due to multiplication of sets of chromosomes (Polyploidy) or it may be due to loss or gain of one or more chromosomes of a diploid set (Aneuploidy) Mutations are also possible due to structural changes in the chromosomes, by deletion, addition, interchange and inversions. Nevertheless, at molecular level, the loss or addition or substitution of nucleotides in a particular segment of genes results in point mutations.
Such a variation at the DNA level results in a change in the function of a cell, so also in its characters. This change is referred to as variation. This variation may be spontaneous or induced, but it is multidirectional. It may by progressive, retrogressive, lethal or non-lethal. It 'may also be small or large, but ultimately the phenotype changes. This accumulation of small or large characters at the genome level leads to speciation and evolution of organisms.